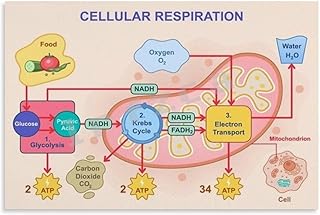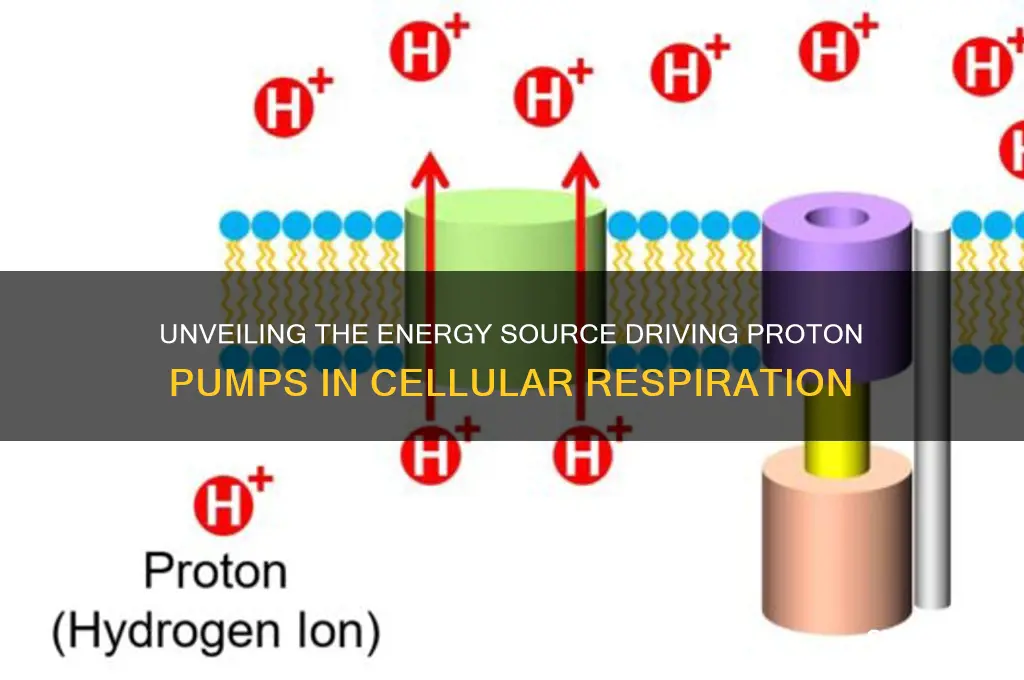
The proton pump, a critical component of cellular respiration, is fueled primarily by the energy derived from the electron transport chain (ETC) in the inner mitochondrial membrane. As electrons are passed along the ETC, they release energy that is captured in the form of ATP through oxidative phosphorylation. Simultaneously, the process creates an electrochemical gradient by pumping protons (H⁺ ions) from the mitochondrial matrix into the intermembrane space. This gradient, known as the proton motive force, serves as the driving force for the proton pump, specifically ATP synthase, to generate ATP. The energy required to maintain this gradient ultimately originates from the oxidation of nutrients like glucose, fatty acids, and amino acids, which are broken down during earlier stages of cellular respiration, such as glycolysis and the citric acid cycle. Thus, the proton pump is fueled by the collective energy harvested from these metabolic pathways.
| Characteristics | Values |
|---|---|
| Energy Source | ATP hydrolysis |
| Mechanism | ATP binds to the proton pump (e.g., ATP synthase), driving conformational changes |
| Proton Movement | Protons (H⁺) are pumped from the mitochondrial matrix to the intermembrane space |
| Location | Inner mitochondrial membrane (in eukaryotes) |
| Role in Cellular Respiration | Establishes an electrochemical gradient (proton motive force) used for ATP synthesis |
| Key Enzyme Involved | ATP synthase (F₀F₁ ATPase) |
| Energy Coupling | Chemiosmosis (coupling of proton flow back into the matrix to ATP synthesis) |
| Fuel for ATP Synthase | Proton gradient (generated by electron transport chain) |
| Direct Fuel for Proton Pump | ATP (not the proton gradient itself) |
| Biological Process | Oxidative phosphorylation |
| Alternative Systems | In some bacteria, light-driven or sodium-driven pumps may operate |
Explore related products






What You'll Learn
- ATP Hydrolysis: Energy from ATP breakdown powers proton pump movement across membranes
- Electron Transport Chain: Electrons drive proton pumping via redox reactions in mitochondria
- Electrochemical Gradient: Proton pumps maintain pH and voltage differences across membranes
- Mitochondrial Structure: Cristae increase surface area for efficient proton pumping
- Proton Motive Force: Gradient fuels ATP synthase and other cellular processes
![]()
ATP Hydrolysis: Energy from ATP breakdown powers proton pump movement across membranes
The proton pump, a critical component in cellular respiration, relies on a precise and efficient energy source to function. This energy comes from the breakdown of adenosine triphosphate (ATP), a molecule often referred to as the "energy currency" of cells. ATP hydrolysis, the process by which ATP is broken down into adenosine diphosphate (ADP) and inorganic phosphate (Pi), releases energy that is harnessed to power the proton pump's movement across membranes. This mechanism is fundamental to maintaining the electrochemical gradient essential for various cellular processes, including ATP synthesis itself.
Consider the step-by-step process of how ATP hydrolysis fuels the proton pump. When ATP binds to the proton pump, it undergoes hydrolysis, releasing approximately 7.3 kcal/mol of free energy. This energy is directly transferred to the pump, enabling it to transport protons against their concentration gradient. For instance, in the electron transport chain of mitochondria, the proton pump (ATP synthase) uses this energy to move protons from the mitochondrial matrix to the intermembrane space. This movement creates a proton gradient, which is later utilized to generate more ATP through chemiosmosis. The efficiency of this process is remarkable, with nearly 100% of the energy from ATP hydrolysis being utilized for proton transport.
From a practical standpoint, understanding ATP hydrolysis and its role in proton pump function has significant implications in fields like medicine and biochemistry. For example, certain drugs, such as oligomycin, inhibit ATP synthase by blocking the proton channel, disrupting ATP production. This knowledge is crucial in developing treatments for conditions like heart failure, where mitochondrial dysfunction plays a role. Additionally, researchers studying aging and metabolic disorders often focus on optimizing ATP hydrolysis efficiency, as declines in this process are linked to cellular energy deficits.
Comparatively, ATP hydrolysis stands out as a highly specialized energy source when contrasted with other cellular energy mechanisms. Unlike glycolysis or the citric acid cycle, which generate ATP, ATP hydrolysis is a direct energy release mechanism. Its specificity in powering proton pumps highlights its unique role in maintaining cellular homeostasis. For instance, while glucose breakdown can produce up to 36 ATP molecules per molecule of glucose, the energy from a single ATP molecule is precisely calibrated to drive the proton pump's mechanical work, ensuring no energy is wasted.
In conclusion, ATP hydrolysis is the linchpin that powers proton pump movement across membranes, a process vital for cellular respiration. Its efficiency, specificity, and practical implications make it a cornerstone of cellular energetics. By breaking down ATP, cells harness the exact amount of energy needed to sustain proton gradients, which in turn drive essential biological processes. Whether in research, medicine, or understanding fundamental biology, the role of ATP hydrolysis in fueling proton pumps remains a critical area of focus.
Why V8 Engines Require Fuel Pump Priming: Essential Insights
You may want to see also
Explore related products






![]()
Electron Transport Chain: Electrons drive proton pumping via redox reactions in mitochondria
The electron transport chain (ETC) is the powerhouse of proton pumping in cellular respiration, a process that hinges on the movement of electrons through a series of redox reactions in the mitochondrial inner membrane. Here’s how it works: electrons derived from NADH and FADH₂, generated during earlier stages of respiration, are passed along protein complexes (I, III, and IV) embedded in the membrane. Each transfer releases energy, which is used to pump protons (H⁺) from the mitochondrial matrix into the intermembrane space. This creates an electrochemical gradient, known as the proton motive force, essential for ATP synthesis. Without the ETC, this gradient—and thus, cellular energy production—would collapse.
Consider the step-by-step mechanism: Complex I accepts electrons from NADH, reducing its cofactors and pumping 4 protons. Electrons then move to ubiquinone (CoQ), a mobile carrier, which delivers them to Complex III. Here, another 4 protons are pumped as electrons are passed to cytochrome *c*. Finally, Complex IV accepts electrons from cytochrome *c* and reduces molecular oxygen to water, pumping 2 more protons. This sequential pumping generates a proton gradient of approximately 4-5 H⁺ per electron pair, depending on the organism. The efficiency of this process is critical; even small disruptions, such as those caused by cyanide (which inhibits Complex IV), halt ATP production entirely.
A comparative analysis highlights the ETC’s elegance: unlike glycolysis or the Krebs cycle, which directly generate ATP, the ETC focuses on creating a proton gradient. This indirect approach is far more efficient, yielding up to 32-34 ATP molecules per glucose molecule, compared to just 2 ATP from glycolysis. The redox reactions driving proton pumping are also highly regulated, with each complex optimized for electron transfer and proton translocation. For instance, Complex IV’s reduction of oxygen to water prevents the formation of reactive oxygen species, a byproduct of incomplete reduction.
Practical implications abound, particularly in medicine. Mitochondrial diseases, often linked to ETC dysfunction, underscore its importance. For example, mutations in Complex I genes can reduce proton pumping efficiency, leading to energy deficits in high-demand tissues like muscle and brain. Treatments targeting these defects, such as coenzyme Q10 supplementation (100-200 mg/day for adults), aim to restore electron flow and proton gradient. Similarly, drugs like metformin, used in diabetes, may enhance ETC activity by increasing AMPK, which upregulates mitochondrial biogenesis.
In conclusion, the ETC’s role in proton pumping is a masterclass in bioenergetics, blending redox chemistry with membrane physics. Its reliance on electron flow, coupled with precise proton translocation, ensures maximal energy extraction from nutrients. Understanding this mechanism not only deepens our appreciation of cellular respiration but also informs therapeutic strategies for metabolic disorders. Whether in a biochemistry lab or a clinical setting, the ETC remains a cornerstone of life’s energy currency.
Why Your Fuel Pump Keeps Blowing: Common Causes and Fixes
You may want to see also
Explore related products






![]()
Electrochemical Gradient: Proton pumps maintain pH and voltage differences across membranes
Proton pumps are the unsung heroes of cellular respiration, tirelessly working to establish and maintain electrochemical gradients across biological membranes. These gradients are not just a byproduct of cellular processes; they are essential for life, driving ATP synthesis and regulating pH levels. At the heart of this mechanism lies the proton pump, a molecular machine fueled by the energy currency of the cell.
The Fuel Behind the Pump: A Metabolic Perspective
The proton pump in cellular respiration, specifically the ATP synthase in mitochondria, is powered by the proton motive force (PMF). This force arises from the movement of protons (H⁺ ions) across the inner mitochondrial membrane. The primary fuel for this process is the electron transport chain (ETC), which uses the energy from NADH and FADH₂, derived from the breakdown of glucose, fatty acids, and amino acids. Each molecule of NADH can generate up to 10 protons, while FADH₂ contributes around 6. These protons are pumped from the mitochondrial matrix into the intermembrane space, creating a concentration gradient. The energy required to pump these protons against their gradient is derived from the redox reactions of the ETC, where electrons are transferred from high-energy molecules to oxygen, releasing energy in the process.
Maintaining the Gradient: A Delicate Balance
Proton pumps not only create the electrochemical gradient but also meticulously maintain it. This gradient consists of two components: a pH difference (due to proton concentration) and a voltage difference (due to the separation of charges). For example, in mitochondria, the pH in the intermembrane space can drop to 6.0, while the matrix remains around 7.8, creating a ΔpH of approximately 1.8 units. Simultaneously, the charge separation generates a membrane potential of about -150 to -180 mV (matrix negative relative to the intermembrane space). This dual gradient is critical for ATP synthase to function, as protons flow back into the matrix through the enzyme, driving the phosphorylation of ADP to ATP.
Practical Implications and Cautions
Understanding the electrochemical gradient is crucial for medical and biochemical applications. For instance, uncouplers like 2,4-dinitrophenol (DNP) disrupt the gradient by allowing protons to leak back into the matrix without generating ATP, leading to increased heat production and potential toxicity. Conversely, inhibitors like oligomycin block ATP synthase, halting ATP production and highlighting the gradient’s central role. In research, maintaining optimal pH and ion concentrations in cell cultures is essential to preserve mitochondrial function. For example, buffers like HEPES (20–25 mM) are commonly used to stabilize pH in experimental conditions, ensuring proton pumps operate efficiently.
Comparative Insights: Beyond Mitochondria
While mitochondria are the prime example, proton pumps and electrochemical gradients are universal in biology. In chloroplasts, light-driven proton pumps generate a gradient used for ATP synthesis during photosynthesis. In bacteria, similar mechanisms power flagellar movement and nutrient transport. Even in acid-loving archaea, proton gradients across the cell membrane drive survival in extreme pH environments. This universality underscores the efficiency and adaptability of proton pumps as a fundamental energy mechanism across life forms.
Takeaway: The Gradient as a Cellular Powerhouse
The electrochemical gradient is more than a byproduct of proton pumping—it’s the linchpin of cellular energy metabolism. By harnessing the energy from redox reactions, proton pumps create a dynamic system that powers ATP synthesis, regulates pH, and sustains life. Whether in mitochondria, chloroplasts, or bacteria, this mechanism exemplifies nature’s ingenuity in converting energy into usable forms. For researchers and clinicians, understanding and manipulating this gradient opens avenues for treating metabolic disorders, optimizing bioenergetics, and engineering synthetic systems inspired by nature’s design.
Resetting Your Ranger's Fuel Pump Shut Off Switch: A Step-by-Step Guide
You may want to see also
Explore related products





![]()
Mitochondrial Structure: Cristae increase surface area for efficient proton pumping
The mitochondrial inner membrane is a bustling hub of energy production, where the proton pump, a key player in cellular respiration, operates with remarkable efficiency. This efficiency is not by chance but by design, thanks to the unique structure of the mitochondria, particularly the cristae. These are inward folds of the inner membrane that significantly increase its surface area, providing more space for the proton pumps, also known as ATP synthase, to work. This structural adaptation is crucial because it allows for a higher density of proton pumps, thereby increasing the rate of ATP production, the cell's primary energy currency.
Imagine a factory assembly line where the number of workstations directly impacts productivity. In the context of mitochondria, the cristae act as additional workstations, enabling more proton pumps to function simultaneously. This is essential during high-energy demands, such as in muscle cells during exercise or in neurons transmitting signals. The increased surface area provided by the cristae ensures that the proton gradient, which drives ATP synthesis, is maintained efficiently, even under stress. For instance, in cardiac muscle cells, which have high energy requirements, the density of cristae is notably higher, reflecting the cell's need for rapid and sustained ATP production.
From a practical standpoint, understanding the role of cristae in proton pumping has implications for health and disease. Conditions such as heart failure or neurodegenerative diseases often involve mitochondrial dysfunction, where the structure of cristae is compromised. For example, in certain mitochondrial diseases, the cristae become disorganized or reduced in number, leading to decreased ATP production. Researchers are exploring therapeutic strategies to enhance cristae structure or function, such as through pharmacological agents that stabilize the inner membrane or genetic therapies targeting mitochondrial proteins. These interventions could potentially restore efficient proton pumping and improve cellular energy levels in affected individuals.
A comparative analysis highlights the evolutionary advantage of cristae in energy-demanding organisms. For example, mammals, with their high metabolic rates, have more complex cristae structures compared to less metabolically active organisms like some invertebrates. This comparison underscores the importance of cristae in adapting to different energy needs across species. Furthermore, studies on hibernating animals show that during periods of reduced metabolic activity, cristae structure can change dynamically, illustrating the flexibility of mitochondrial architecture in response to physiological demands.
In conclusion, the cristae of the mitochondrial inner membrane are not merely structural features but essential components that optimize proton pumping efficiency. Their role in increasing surface area for ATP synthase highlights the intricate relationship between cellular structure and function. By focusing on cristae, researchers and clinicians can develop targeted interventions to address energy deficiencies in various diseases, ultimately improving health outcomes. Whether in the context of high-performance athletics or managing chronic illnesses, the cristae remind us of the remarkable adaptability and efficiency of cellular respiration.
Locating the TSX Fuel Pump Relay: A Comprehensive Guide
You may want to see also
Explore related products






![]()
Proton Motive Force: Gradient fuels ATP synthase and other cellular processes
The proton motive force (PMF) is a critical energy currency in cellular respiration, driving ATP synthesis and other essential processes. This force arises from the electrochemical gradient of protons (H⁺) across the inner mitochondrial membrane, created by the electron transport chain (ETC). As electrons pass through the ETC, they are coupled to the pumping of protons from the mitochondrial matrix into the intermembrane space, generating a pH difference and an electric potential. This gradient, the PMF, is the fuel that powers ATP synthase, the enzyme responsible for phosphorylating ADP to ATP.
Mechanics of PMF Utilization:
ATP synthase acts as a molecular turbine, harnessing the energy from the proton gradient to perform mechanical work. Protons flow back into the matrix through a channel in ATP synthase, a process known as chemiosmosis. This movement drives the rotation of a central stalk within the enzyme, which in turn induces conformational changes in the catalytic sites, enabling ATP production. Remarkably, this mechanism is conserved across species, from bacteria to humans, underscoring its evolutionary significance. The efficiency of ATP synthase is near-perfect, converting approximately 90% of the PMF energy into chemical energy.
Beyond ATP Synthesis: PMF’s Versatile Role:
While ATP synthase is the most prominent consumer of PMF, this gradient also fuels other cellular processes. For instance, in bacteria, PMF drives the active transport of nutrients across the cell membrane, ensuring survival in nutrient-poor environments. In eukaryotic cells, PMF is involved in protein import into mitochondria, maintaining organelle function. Additionally, PMF regulates ion homeostasis, influencing processes like muscle contraction and neuronal signaling. This versatility highlights the PMF as a central hub of cellular energy management.
Practical Implications and Optimization:
Understanding PMF has practical applications in medicine and biotechnology. For example, uncouplers like 2,4-dinitrophenol dissipate the proton gradient, increasing metabolic rate but also causing energy wastage, which has been explored in weight loss treatments (though with significant risks). Conversely, inhibitors of the ETC, such as rotenone, block proton pumping, disrupting PMF and ATP production—a mechanism exploited in pesticides. To optimize PMF-dependent processes, researchers focus on enhancing mitochondrial health through diet (e.g., coenzyme Q10 supplementation) and exercise, which boost ETC efficiency. For older adults (ages 65+), maintaining PMF integrity is crucial, as mitochondrial dysfunction is linked to aging and diseases like Parkinson’s.
Comparative Perspective: PMF Across Organisms:
The reliance on PMF varies across life forms. In plants, PMF generated in chloroplasts during photosynthesis supports ATP synthesis and NADPH production for carbon fixation. In contrast, some anaerobic bacteria use alternative gradients, such as sodium ions, to drive ATP synthesis, showcasing nature’s adaptability. However, the proton gradient remains the most widespread mechanism due to protons’ abundance and mobility. This comparative view emphasizes the PMF’s role as a unifying principle in bioenergetics, bridging diverse organisms and metabolic pathways.
Understanding the Mechanism: How Injection Pumps Shut Off Fuel Supply
You may want to see also
Frequently asked questions
The proton pump in cellular respiration is fueled by the energy released from the electron transport chain (ETC). As electrons are passed through the ETC, energy is harnessed to pump protons (H⁺ ions) from the mitochondrial matrix into the intermembrane space, creating an electrochemical gradient.
The energy for proton pumping comes from the redox reactions occurring in the electron transport chain. Electrons derived from NADH and FADH₂ are transferred through protein complexes, releasing energy that is used to actively transport protons across the inner mitochondrial membrane.
The proton pump maintains the proton gradient by coupling the exergonic (energy-releasing) process of electron transfer in the ETC to the endergonic (energy-requiring) process of proton translocation. This creates a concentration and electrical potential difference across the membrane, which is later used by ATP synthase to generate ATP.