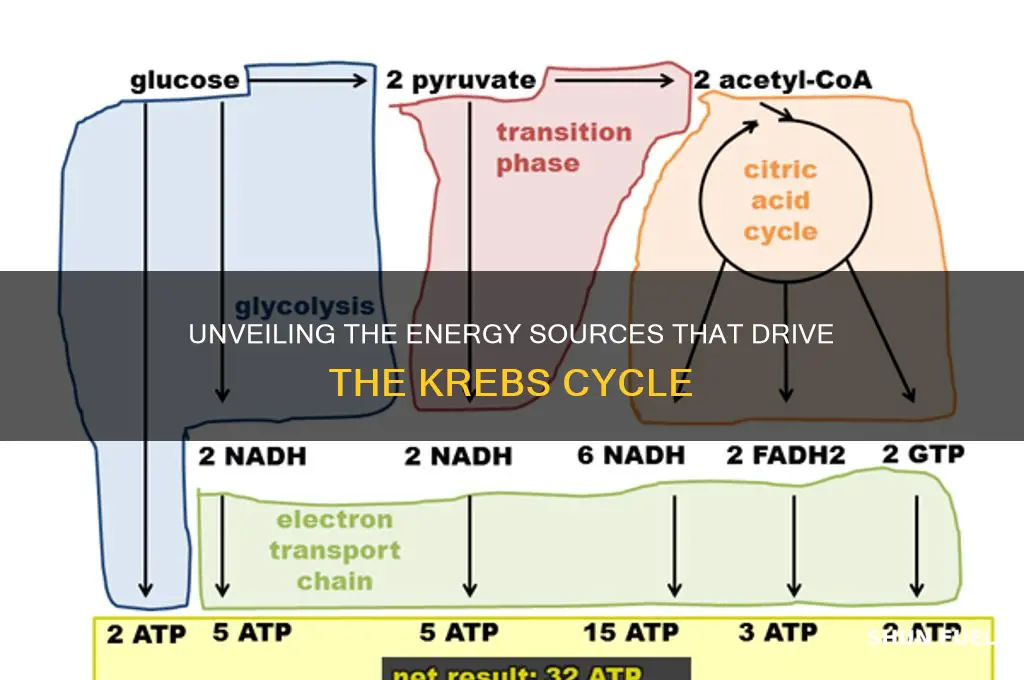
The Krebs cycle, also known as the citric acid cycle, is a central metabolic pathway that generates energy in the form of ATP by oxidizing acetyl-CoA derived from carbohydrates, fats, and proteins. This cycle is fueled primarily by acetyl-CoA, which is produced during the breakdown of glucose (via glycolysis), fatty acids (via beta-oxidation), and amino acids (via various catabolic pathways). Once inside the mitochondria, acetyl-CoA combines with oxaloacetate to form citrate, initiating a series of enzymatic reactions that release high-energy molecules like NADH and FADH₂, which are then used in the electron transport chain to produce ATP. Thus, the Krebs cycle serves as a critical hub for energy production, linking the metabolism of diverse macronutrients to cellular respiration.
| Characteristics | Values |
|---|---|
| Primary Fuel | Acetyl-CoA (derived from pyruvate, fatty acids, or amino acids) |
| Key Substrates | Pyruvate (from glycolysis), fatty acids (from beta-oxidation), amino acids (from protein metabolism) |
| Starting Molecule | Citrate (formed by combining acetyl-CoA with oxaloacetate) |
| Energy Carriers Produced | NADH, FADH2, ATP (or GTP) |
| CO2 Release | 2 molecules of CO2 per acetyl-CoA |
| Redox Reactions | Oxidation of acetyl-CoA and reduction of NAD+ and FAD to NADH and FADH2 |
| Regeneration of Oxaloacetate | Ensures the cycle continues by reforming the starting molecule |
| Location | Mitochondrial matrix in eukaryotes |
| Steps | 8 enzymatic steps involving decarboxylation, dehydrogenation, and substrate-level phosphorylation |
| Role in Metabolism | Central hub for carbohydrate, fat, and protein metabolism |
| Oxygen Requirement | Aerobic process (requires oxygen for electron transport chain) |
| ATP Yield per Glucose | ~10 ATP (combined with glycolysis and oxidative phosphorylation) |
Explore related products






What You'll Learn
- Pyruvate Oxidation: Converts pyruvate to acetyl-CoA, entering the Krebs cycle for energy production
- Acetyl-CoA Formation: Derived from fats, carbs, proteins, feeding into the Krebs cycle
- NADH and FADH2: Electron carriers generated, fueling oxidative phosphorylation for ATP synthesis
- Citric Acid Formation: Combines acetyl-CoA with oxaloacetate, initiating the cycle reactions
- Substrate Availability: Depends on metabolic pathways like glycolysis, beta-oxidation, amino acid breakdown
![]()
Pyruvate Oxidation: Converts pyruvate to acetyl-CoA, entering the Krebs cycle for energy production
Pyruvate oxidation stands as the critical gateway to the Krebs cycle, transforming pyruvate—a byproduct of glycolysis—into acetyl-CoA, the molecule that fuels this central metabolic pathway. This process occurs in the mitochondrial matrix and involves a series of enzymatic reactions that not only generate acetyl-CoA but also produce a molecule of NADH, an essential electron carrier in the electron transport chain. Without pyruvate oxidation, the Krebs cycle would lack its primary substrate, halting energy production in aerobic organisms.
Consider the step-by-step mechanism of pyruvate oxidation. First, pyruvate dehydrogenase (PDH) decarboxylates pyruvate, releasing carbon dioxide and forming a hydroxyethyl group. This group is then oxidized, transferring electrons to NAD+ to form NADH. Finally, the oxidized group is bound to coenzyme A, yielding acetyl-CoA. This process is highly regulated, with PDH activity influenced by factors like energy demand, calcium levels, and the availability of NAD+. For instance, high NADH levels inhibit PDH, preventing overproduction of acetyl-CoA when energy stores are sufficient.
From a practical standpoint, understanding pyruvate oxidation is crucial for optimizing metabolic health. Athletes and fitness enthusiasts can enhance this process by maintaining adequate levels of thiamine (vitamin B1), a cofactor for PDH. Additionally, moderate carbohydrate intake ensures a steady supply of pyruvate, while avoiding excessive sugar consumption prevents metabolic overload. For individuals with metabolic disorders, such as diabetes, supporting pyruvate oxidation through diet and lifestyle modifications can improve energy efficiency and reduce complications.
Comparatively, pyruvate oxidation differs from anaerobic fermentation, which occurs in oxygen-deprived conditions. In fermentation, pyruvate is converted to lactate or ethanol, bypassing the Krebs cycle and producing far less ATP. This highlights the efficiency of aerobic metabolism, where pyruvate oxidation and the Krebs cycle together generate up to 36 ATP molecules per glucose molecule, compared to just 2 ATP in anaerobic pathways. This distinction underscores the evolutionary advantage of aerobic respiration in complex organisms.
In conclusion, pyruvate oxidation is not merely a biochemical reaction but a linchpin of energy metabolism. By converting pyruvate to acetyl-CoA, it ensures the Krebs cycle’s continuity, driving ATP production and sustaining life. Whether in the context of athletic performance, metabolic health, or evolutionary biology, this process exemplifies the elegance and efficiency of cellular energy systems.
Raw Fuel Emissions: Environmental Impact and Harmful Pollutants Explained
You may want to see also
Explore related products


$9.49




![]()
Acetyl-CoA Formation: Derived from fats, carbs, proteins, feeding into the Krebs cycle
The Krebs cycle, a cornerstone of cellular energy production, relies on a single molecular currency: acetyl-CoA. This two-carbon compound acts as the universal fuel, regardless of whether the body burns fats, carbohydrates, or proteins for energy. Understanding how these diverse macronutrients converge into acetyl-CoA reveals the elegant efficiency of our metabolic machinery.
Fat, a concentrated energy source, undergoes beta-oxidation, a process akin to molecular dismantling. Each fatty acid chain is systematically broken down, releasing acetyl-CoA units like energy-rich bricks. This process is particularly crucial during prolonged fasting or low-carbohydrate diets, when fats become the primary energy substrate. Interestingly, the longer the fatty acid chain, the more acetyl-CoA molecules are generated, highlighting the efficiency of fat as a fuel source.
Carbohydrates, our body's preferred energy source, follow a different path. Glucose, the simplest carbohydrate, is first broken down through glycolysis, yielding pyruvate. This pyruvate then enters the mitochondria, where it's converted to acetyl-CoA through a process called pyruvate dehydrogenase complex (PDC) reaction. This step is a critical juncture, as it links carbohydrate metabolism directly to the Krebs cycle. It's worth noting that the PDC reaction is highly regulated, ensuring a steady supply of acetyl-CoA without overwhelming the cycle.
A less direct but equally important contributor is protein. Amino acids, the building blocks of proteins, can be deaminated, removing their nitrogen-containing groups. The remaining carbon skeletons can then be converted into intermediates of the Krebs cycle, ultimately feeding into acetyl-CoA production. This pathway is particularly active during periods of protein breakdown, such as intense exercise or starvation. However, excessive reliance on protein for energy can lead to muscle wasting, underscoring the importance of a balanced diet.
Understanding acetyl-CoA formation from diverse sources has practical implications. For athletes, optimizing carbohydrate intake before and during exercise ensures a readily available source of acetyl-CoA for peak performance. Individuals on low-carb diets need to ensure adequate fat intake to sustain acetyl-CoA production and prevent energy deficits. Moreover, recognizing the potential drawbacks of excessive protein breakdown for energy highlights the importance of sufficient carbohydrate and fat intake for overall metabolic health. By appreciating the multifaceted origins of acetyl-CoA, we gain a deeper understanding of how our bodies adapt to different dietary conditions and fuel our cellular engines.
Does RAM Monitor Air-Fuel Ratio? Understanding Vehicle Performance Metrics
You may want to see also
Explore related products


![Essencea Citric Acid 5LB Pure Bulk Ingredients | Non-GMO | 100% Pure Citric Acid Powder [Packaging May Vary]](https://m.media-amazon.com/images/I/51O7Lk96ljL._AC_UL320_.jpg)



![]()
NADH and FADH2: Electron carriers generated, fueling oxidative phosphorylation for ATP synthesis
The Krebs cycle, a central metabolic pathway, relies on electron carriers to shuttle energy derived from nutrients to the electron transport chain (ETC), ultimately driving ATP synthesis. Among these carriers, NADH and FADH₂ play pivotal roles, each generated through distinct steps in the cycle. NADH is produced during the oxidation of isocitrate to α-ketoglutarate and α-ketoglutarate to succinyl-CoA, while FADH₂ arises from the conversion of succinate to fumarate. These molecules act as energy-rich intermediaries, ferrying electrons from the Krebs cycle to the ETC, where their potential is harnessed to generate ATP via oxidative phosphorylation.
Consider the efficiency of these carriers: NADH yields approximately 2.5 ATP molecules per molecule, whereas FADH₂ produces around 1.5 ATP molecules. This disparity stems from the differing entry points of their electrons into the ETC. NADH transfers electrons at a higher energy level (Complex I), allowing for more ATP production, while FADH₂ enters at a lower energy level (Complex II), resulting in fewer ATP molecules. Understanding this distinction highlights the Krebs cycle’s strategic use of these carriers to maximize energy extraction from acetyl-CoA, the cycle’s primary substrate.
Practical implications of NADH and FADH₂ extend beyond cellular metabolism. For instance, in exercise physiology, increased demand for ATP elevates the production of these carriers, emphasizing the importance of aerobic capacity. Athletes can enhance NADH and FADH₂ generation by consuming carbohydrate-rich diets, ensuring sufficient glucose for glycolysis and subsequent entry into the Krebs cycle. Conversely, fasting or low-carb diets may reduce substrate availability, limiting NADH and FADH₂ production and, consequently, ATP synthesis.
A comparative analysis reveals the interplay between NADH, FADH₂, and other metabolic pathways. While NADH is also generated in glycolysis, its production in the Krebs cycle is more ATP-efficient due to the cycle’s ability to fully oxidize acetyl-CoA. FADH₂, though less ATP-yielding, complements NADH by ensuring a continuous electron flow, preventing bottlenecks in the ETC. This synergy underscores the Krebs cycle’s role as a metabolic hub, integrating multiple pathways to sustain cellular energy demands.
In summary, NADH and FADH₂ are indispensable electron carriers that bridge the Krebs cycle and oxidative phosphorylation, converting the energy embedded in nutrients into usable ATP. Their distinct roles, efficiencies, and interactions with metabolic pathways make them critical targets for optimizing energy production, whether in cellular biology, athletic performance, or dietary strategies. By understanding their mechanisms, one can appreciate the elegance of the Krebs cycle and its centrality in fueling life’s processes.
Do Fuel Max Tires Really Save Gas? A Comprehensive Review
You may want to see also
Explore related products






![]()
Citric Acid Formation: Combines acetyl-CoA with oxaloacetate, initiating the cycle reactions
The Krebs cycle, a cornerstone of cellular respiration, begins with a pivotal reaction: the combination of acetyl-CoA and oxaloacetate to form citrate. This step, catalyzed by citrate synthase, is not merely a chemical transformation but the ignition point for a series of reactions that generate ATP, NADH, and FADH₂, essential for energy production. Acetyl-CoA, derived from the breakdown of carbohydrates, fats, and proteins, acts as the primary fuel for this process, while oxaloacetate serves as the catalytic substrate, regenerating at the cycle’s end to sustain continuous operation.
To visualize this process, imagine acetyl-CoA as a molecular key and oxaloacetate as the lock. When they combine, the resulting citrate molecule unlocks the Krebs cycle’s energy-harvesting potential. This reaction is highly regulated, as the cell must balance the availability of acetyl-CoA with the demand for energy. For instance, during fasting, increased fatty acid breakdown elevates acetyl-CoA levels, accelerating the cycle. Conversely, high carbohydrate intake boosts pyruvate production, which is converted to acetyl-CoA via pyruvate dehydrogenase, further fueling the cycle.
Practical considerations for optimizing this reaction include dietary choices. Consuming a balanced diet rich in complex carbohydrates, healthy fats, and lean proteins ensures a steady supply of acetyl-CoA precursors. For athletes or individuals with high energy demands, timing nutrient intake around physical activity can enhance acetyl-CoA availability, thereby maximizing ATP production. However, excessive calorie intake, particularly from simple sugars, can overwhelm the cycle, leading to incomplete oxidation and increased oxidative stress.
A cautionary note: disruptions in citrate formation, such as deficiencies in B vitamins (cofactors for pyruvate dehydrogenase) or genetic disorders affecting acetyl-CoA synthesis, can impair the Krebs cycle. For example, individuals with pyruvate dehydrogenase deficiency may experience lactic acidosis and neurological symptoms due to reduced acetyl-CoA production. Supplementation with thiamine (B1), riboflavin (B2), and niacin (B3) can support enzyme function, but dosages should be tailored to age and health status—typically 1.1–1.3 mg/day of thiamine for adults and 1.0–1.6 mg/day of riboflavin, under professional guidance.
In conclusion, citrate formation is the linchpin of the Krebs cycle, bridging nutrient breakdown and energy generation. By understanding its mechanisms and dependencies, individuals can strategically support this process through diet, supplementation, and lifestyle choices. Whether for athletic performance, metabolic health, or disease prevention, optimizing acetyl-CoA and oxaloacetate interaction offers a tangible pathway to enhance cellular energy production.
Mastering Fiber Fuel Combustion: Efficient Techniques for Optimal Energy Output
You may want to see also
Explore related products






![]()
Substrate Availability: Depends on metabolic pathways like glycolysis, beta-oxidation, amino acid breakdown
The Krebs cycle, also known as the citric acid cycle, is a central metabolic pathway that generates energy in the form of ATP. However, its efficiency hinges on the availability of specific substrates, which are derived from various metabolic pathways. Understanding these pathways—glycolysis, beta-oxidation, and amino acid breakdown—is crucial for optimizing energy production and metabolic health.
Glycolysis: The Carbohydrate Connection
Glycolysis is the initial step in breaking down glucose, a primary energy source. This pathway converts one molecule of glucose into two molecules of pyruvate, which can then enter the Krebs cycle as acetyl-CoA. For example, during high-intensity exercise, muscles rely heavily on glycolysis to fuel the Krebs cycle rapidly. However, excessive carbohydrate intake without corresponding energy expenditure can lead to pyruvate overload, potentially disrupting metabolic balance. To optimize substrate availability, aim for a balanced carbohydrate intake, especially for active individuals, with a focus on complex carbs like whole grains and vegetables.
Beta-Oxidation: Tapping into Fat Reserves
When carbohydrate availability is low, the body shifts to fats as the primary fuel source. Beta-oxidation breaks down fatty acids into acetyl-CoA, which directly feeds the Krebs cycle. This pathway is particularly active during fasting, low-carb diets, or prolonged moderate-intensity exercise. For instance, athletes engaging in endurance activities can enhance fat utilization by incorporating medium-chain triglycerides (MCTs) into their diet, as MCTs are more readily oxidized. However, excessive reliance on fats without adequate carbohydrate intake can lead to ketosis, which may not be optimal for all individuals. A practical tip is to include healthy fats like avocados, nuts, and olive oil while monitoring energy levels and metabolic markers.
Amino Acid Breakdown: The Protein Contribution
Amino acids, derived from protein breakdown, can also fuel the Krebs cycle via gluconeogenesis and direct conversion to intermediates like alpha-ketoglutarate or oxaloacetate. This pathway is particularly important during starvation or extreme dieting when other fuel sources are depleted. For example, branched-chain amino acids (BCAAs) like leucine can be catabolized to produce acetyl-CoA. However, excessive protein breakdown for energy can lead to muscle wasting and metabolic stress. To maintain a balance, ensure adequate protein intake (1.2–2.0 g/kg body weight daily) while avoiding prolonged calorie deficits. Pairing protein with resistance training can further preserve muscle mass and optimize substrate availability.
Practical Takeaways for Substrate Optimization
To maximize Krebs cycle efficiency, diversify your fuel sources by balancing carbohydrates, fats, and proteins. For instance, a pre-workout meal combining carbs (e.g., oats), fats (e.g., nuts), and protein (e.g., Greek yogurt) can provide sustained energy. Monitor metabolic responses through biomarkers like blood glucose and ketone levels to fine-tune intake. Additionally, consider timing nutrient intake around activity levels—carbohydrates pre-exercise for glycolytic support and fats post-exercise for recovery. By strategically leveraging glycolysis, beta-oxidation, and amino acid breakdown, you can ensure a steady supply of substrates to fuel the Krebs cycle and meet energy demands effectively.
Understanding Jet Fuel Weight: Key Factors and Calculations Explained
You may want to see also
Frequently asked questions
The primary fuel for the Krebs cycle is acetyl-CoA, which is derived from the breakdown of carbohydrates, fats, and proteins.
Glucose fuels the Krebs cycle by first being broken down into pyruvate through glycolysis, which is then converted to acetyl-CoA, the direct substrate for the cycle.
Yes, fatty acids can fuel the Krebs cycle through beta-oxidation, which produces acetyl-CoA that enters the cycle.
Amino acids can be deaminated and converted into intermediates of the Krebs cycle, such as alpha-ketoglutarate or oxaloacetate, which then fuel the cycle.
No, the Krebs cycle does not use oxygen directly as fuel. Oxygen is required for the electron transport chain, which follows the Krebs cycle, but not as a substrate for the cycle itself.