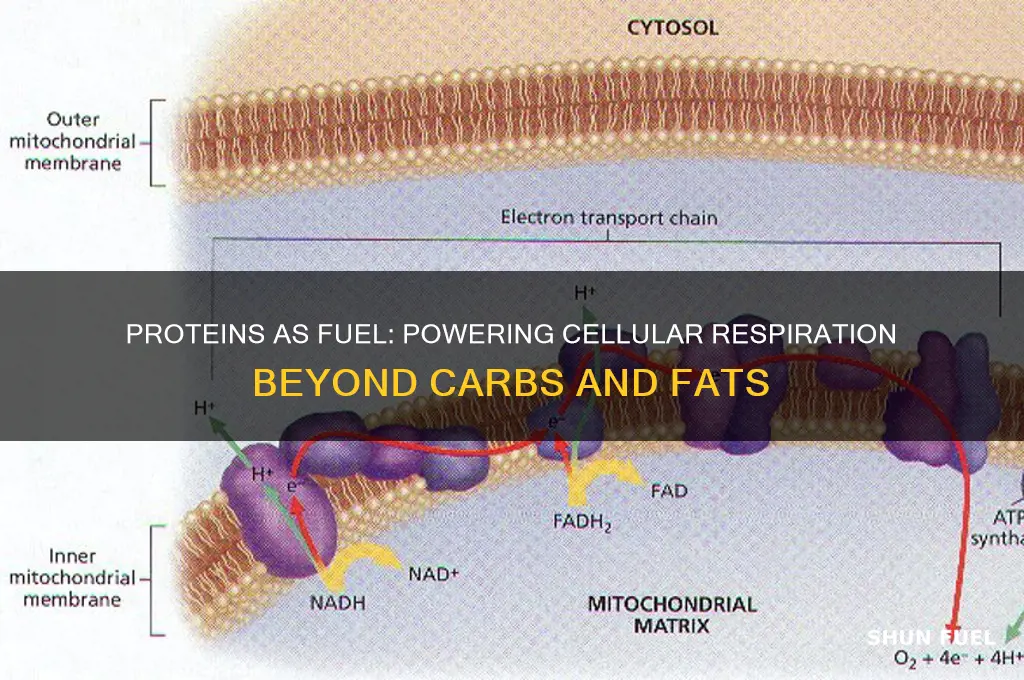
Proteins, while primarily known for their structural and enzymatic roles in the body, can also serve as a source of energy to fuel cellular respiration under specific conditions. Typically, carbohydrates and fats are the preferred substrates for ATP production, but when these sources are depleted, such as during prolonged fasting or intense physical activity, the body can break down proteins into amino acids. These amino acids are then deaminated to remove the nitrogen-containing groups, converting them into molecules like pyruvate or acetyl-CoA, which can enter the citric acid cycle and oxidative phosphorylation pathways. Although this process is less efficient than using carbohydrates or fats, it provides a crucial energy reserve, ensuring cellular functions continue when other fuel sources are scarce. However, reliance on protein for energy can lead to muscle wasting and other metabolic imbalances, highlighting the importance of maintaining a balanced diet to preserve protein’s primary functions.
| Characteristics | Values |
|---|---|
| Primary Energy Source | Proteins are not the primary energy source for cellular respiration; carbohydrates (glucose) and fats are preferred. |
| Usage in Energy Production | Proteins can be used as a fuel source for cellular respiration, but only under specific conditions, such as during prolonged starvation or intense exercise when carbohydrate and fat reserves are depleted. |
| Metabolic Pathway | Proteins are broken down into amino acids, which can be deaminated to remove the nitrogen-containing part. The remaining carbon skeleton enters the citric acid cycle (Krebs cycle) for ATP production. |
| Efficiency | Less efficient compared to carbohydrates and fats. Only about 4 kcal/g of energy is derived from proteins, versus 9 kcal/g for fats and 4 kcal/g for carbohydrates. |
| Byproduct Formation | Produces ammonia (NH₃) as a byproduct, which is toxic and must be converted to urea in the liver for excretion. |
| Gluconeogenesis | Some amino acids can be converted into glucose via gluconeogenesis, providing an indirect source of energy for cellular respiration. |
| Muscle Breakdown | During prolonged fasting or starvation, muscle proteins may be broken down to provide amino acids for energy, leading to muscle wasting. |
| Regulation | Tightly regulated by hormones like glucagon and cortisol, which increase protein breakdown during stress or fasting. |
| Role in Normal Conditions | Under normal conditions, proteins contribute less than 5% of total energy needs, as the body prioritizes preserving protein for structural and functional roles. |
| Health Implications | Excessive reliance on proteins for energy can lead to kidney stress, electrolyte imbalances, and increased risk of osteoporosis due to calcium loss. |
Explore related products






What You'll Learn
- Protein breakdown into amino acids for energy production
- Role of gluconeogenesis in converting amino acids to glucose
- Ketogenesis from amino acid metabolism during prolonged fasting
- Impact of protein oxidation on ATP generation in mitochondria
- Comparison of protein efficiency versus carbohydrates and fats in respiration
![]()
Protein breakdown into amino acids for energy production
Proteins, while primarily known for their structural and functional roles in the body, can indeed be utilized as a source of energy through a process that involves their breakdown into amino acids. This mechanism is particularly important during periods of prolonged starvation, intense exercise, or when carbohydrate and fat reserves are depleted. The process begins with the digestion of proteins in the gastrointestinal tract, where enzymes such as pepsin and trypsin break down dietary proteins into smaller peptides and individual amino acids. These amino acids are then absorbed into the bloodstream and transported to the liver, where the majority of protein metabolism occurs.
Once in the liver, amino acids undergo a series of metabolic pathways to be converted into energy. The first step involves the removal of the amino group (-NH₂) from the amino acid, a process known as deamination. This reaction is catalyzed by enzymes such as transaminases and oxidases, producing ammonia (NH₃) as a byproduct. Ammonia is toxic and must be converted into a less harmful substance, urea, through the urea cycle before being excreted by the kidneys. The remaining carbon skeleton of the amino acid can then enter central metabolic pathways to generate energy.
The carbon skeletons derived from amino acids are funneled into either the tricarboxylic acid (TCA) cycle or gluconeogenesis, depending on the body's energy needs. For instance, glucogenic amino acids can be converted into glucose through gluconeogenesis, providing a source of energy for tissues that rely on glucose, such as the brain. Ketogenic amino acids, on the other hand, are converted into ketone bodies or acetyl-CoA, which can be used directly in the TCA cycle to produce ATP. This flexibility allows the body to utilize amino acids as a versatile energy source when other fuels are scarce.
It is important to note that protein breakdown for energy is not the body's preferred method of energy production. Carbohydrates and fats are typically the primary fuels for cellular respiration due to their higher efficiency and the lower metabolic cost of their breakdown. However, during conditions of energy deficit, the body prioritizes survival, and proteins become a critical energy reserve. This process is tightly regulated to prevent excessive muscle wasting and maintain essential protein functions.
In summary, proteins can be broken down into amino acids and utilized for energy production through deamination, gluconeogenesis, and the TCA cycle. While this pathway is not the body's first choice for energy, it serves as a vital backup system during times of metabolic stress. Understanding this process highlights the adaptability of human metabolism and the multifaceted role of proteins in maintaining cellular function and energy homeostasis.
Bad Fuel Impact: Does It Lead to Excessive Oil Consumption?
You may want to see also
Explore related products






![]()
Role of gluconeogenesis in converting amino acids to glucose
Proteins can indeed be used to fuel cellular respiration, but not directly. Unlike carbohydrates and fats, proteins are not the preferred energy source for cells under normal conditions. However, during prolonged fasting, intense exercise, or certain metabolic states, amino acids derived from protein breakdown can be converted into glucose through a process called gluconeogenesis. This pathway is crucial for maintaining blood glucose levels and providing energy to tissues that rely on glucose, such as the brain and red blood cells. Gluconeogenesis primarily occurs in the liver, with minor contributions from the kidneys, and it ensures that the body has a steady supply of glucose even when dietary intake is insufficient.
The role of gluconeogenesis in converting amino acids to glucose begins with the breakdown of proteins into their constituent amino acids. Not all amino acids can be used for gluconeogenesis; only glucogenic amino acids are suitable for this process. These amino acids, such as alanine, glutamine, and glycine, can be converted into intermediates of the citric acid cycle (e.g., oxaloacetate, α-ketoglutarate) or directly into pyruvate. Once these intermediates are formed, they enter the gluconeogenic pathway, which involves a series of enzymatic reactions that reverse the glycolytic process. Key enzymes in gluconeogenesis include phosphoenolpyruvate carboxykinase (PEPCK) and fructose-1,6-bisphosphatase (FBPase), which catalyze irreversible steps that bypass the energy-releasing steps of glycolysis.
The conversion of amino acids to glucose via gluconeogenesis is energetically costly, requiring six molecules of ATP per molecule of glucose produced, compared to the two molecules of ATP yielded from glycolysis. Despite this inefficiency, gluconeogenesis is essential for survival during periods of carbohydrate deprivation. For example, during prolonged fasting, muscle protein is broken down to release alanine, which is transported to the liver. There, alanine is converted to pyruvate, which serves as a substrate for gluconeogenesis. This process not only provides glucose for the brain and other glucose-dependent tissues but also helps prevent excessive protein catabolism by recycling amino acid carbons into a more usable energy source.
Gluconeogenesis is tightly regulated to ensure that it occurs only when necessary. Hormones such as glucagon, cortisol, and adrenaline stimulate gluconeogenesis by activating key enzymes and promoting the breakdown of proteins and glycerol (from fats) to provide substrates. Insulin, on the other hand, inhibits gluconeogenesis by downregulating PEPCK and FBPase, as sufficient glucose is already available from dietary sources. This hormonal regulation ensures that gluconeogenesis is activated only when blood glucose levels drop, such as during fasting or intense physical activity.
In summary, gluconeogenesis plays a vital role in converting amino acids derived from proteins into glucose, thereby allowing proteins to indirectly fuel cellular respiration. While this pathway is not the primary means of energy production under normal conditions, it becomes critical during states of carbohydrate depletion. By recycling amino acid carbons into glucose, gluconeogenesis supports the energy needs of essential tissues and maintains metabolic homeostasis. Understanding this process highlights the flexibility of human metabolism and the interconnectedness of macronutrient utilization in energy production.
Can Ingles Gift Cards Boost Your Fuel Points Earnings?
You may want to see also
Explore related products






![]()
Ketogenesis from amino acid metabolism during prolonged fasting
During prolonged fasting, the body undergoes significant metabolic adaptations to maintain energy homeostasis. While glucose is the primary fuel for cellular respiration under normal conditions, its availability diminishes during fasting, prompting the body to utilize alternative energy sources. One such source is amino acids derived from protein breakdown. Although proteins are not the preferred fuel for cellular respiration due to their primary role in structural and functional processes, they can be catabolized to provide energy when carbohydrate stores are depleted. This process involves the deamination of amino acids, where the nitrogen-containing portion is removed, and the carbon skeleton is converted into intermediates that enter the citric acid cycle (Krebs cycle) for ATP production. However, prolonged reliance on amino acids for energy is unsustainable due to the risk of muscle wasting and impaired protein function.
Ketogenesis emerges as a critical metabolic pathway during prolonged fasting to spare protein and provide an alternative energy source. When carbohydrate availability is low, the liver increases the production of ketone bodies (acetoacetate, β-hydroxybutyrate, and acetone) from fatty acids. However, amino acid metabolism also plays a role in ketogenesis, particularly through the contribution of specific amino acids. For instance, the catabolism of certain amino acids, such as leucine and lysine, yields acetyl-CoA, a key substrate for ketone body synthesis. Additionally, the carbon skeletons of other amino acids can be converted into glucose via gluconeogenesis, indirectly supporting ketogenesis by maintaining the supply of oxaloacetate in the liver. This interplay between amino acid metabolism and ketogenesis ensures that proteins are not excessively degraded for energy while providing a sustainable fuel source for tissues like the brain and muscles.
The process of ketogenesis from amino acid metabolism is tightly regulated to balance energy needs with protein preservation. During prolonged fasting, the hormone glucagon increases, promoting the breakdown of proteins into amino acids in skeletal muscle. These amino acids are then transported to the liver, where they undergo deamination and transamination. The resulting carbon skeletons can either enter the citric acid cycle directly or be converted into glucose via gluconeogenesis. Simultaneously, the increased availability of acetyl-CoA from both fatty acid oxidation and amino acid catabolism drives ketogenesis. This dual utilization of amino acids—both as a direct energy source and as a contributor to ketone body production—highlights their role in sustaining cellular respiration during fasting while minimizing muscle protein loss.
Ketone bodies produced during prolonged fasting become the primary fuel for many tissues, including the brain, which can derive up to 70% of its energy from ketones. This shift reduces the reliance on glucose and amino acids for energy, effectively sparing protein. The brain’s ability to utilize ketones is particularly important, as it cannot directly metabolize fatty acids. Meanwhile, other tissues, such as skeletal muscle, increase their uptake of ketone bodies, further decreasing the demand for amino acid oxidation. This metabolic flexibility ensures that cellular respiration continues efficiently even in the absence of dietary carbohydrate and protein intake.
In summary, ketogenesis from amino acid metabolism during prolonged fasting is a vital adaptive mechanism that supports cellular respiration while preserving protein integrity. Amino acids contribute to ketogenesis indirectly through the production of acetyl-CoA and by maintaining gluconeogenic substrates. This process, coupled with the increased utilization of ketone bodies by peripheral tissues, ensures energy homeostasis without excessive protein catabolism. Understanding this interplay between amino acid metabolism and ketogenesis provides insights into the body’s remarkable ability to sustain function during extended periods of nutrient deprivation.
Can Truck Drivers Deduct Fuel Costs on Their Taxes?
You may want to see also
Explore related products






![]()
Impact of protein oxidation on ATP generation in mitochondria
Proteins can indeed be used to fuel cellular respiration, although they are not the preferred energy source compared to carbohydrates and fats. Under conditions of carbohydrate and fat depletion, such as prolonged fasting or intense exercise, proteins are broken down into amino acids, which can enter the citric acid cycle (Krebs cycle) after deamination and conversion to acetyl-CoA or other intermediates. This process allows proteins to contribute to ATP production, albeit at a lower efficiency compared to glucose or fatty acids. However, the oxidation of proteins for energy has significant implications, particularly in the mitochondria, where ATP generation occurs via oxidative phosphorylation.
Protein oxidation, a process where proteins undergo oxidative modification due to reactive oxygen species (ROS), can directly impact mitochondrial function and ATP generation. Mitochondria are both the primary site of ATP production and a major source of ROS, creating a delicate balance between energy generation and oxidative stress. When proteins in the mitochondrial electron transport chain (ETC) or other key enzymes are oxidized, their structure and function can be compromised. For instance, oxidation of ETC complexes (e.g., Complex I or IV) reduces their efficiency in transferring electrons, leading to decreased proton pumping across the inner mitochondrial membrane. This disruption diminishes the proton gradient, which is essential for ATP synthase to generate ATP, ultimately reducing cellular energy output.
The impact of protein oxidation extends beyond the ETC to other mitochondrial proteins involved in ATP production. Enzymes in the citric acid cycle, such as aconitase and α-ketoglutarate dehydrogenase, are particularly susceptible to oxidation due to their iron-sulfur clusters. Oxidative modification of these enzymes impairs their catalytic activity, slowing the cycle and reducing the availability of NADH and FADH2, which are critical electron donors for the ETC. This cascade of events further limits ATP synthesis and exacerbates energy deficiency in cells reliant on protein oxidation for fuel.
Additionally, protein oxidation can trigger mitochondrial dysfunction through mechanisms beyond direct enzyme inactivation. Oxidized proteins may aggregate or misfold, leading to mitochondrial protein quality control issues. The accumulation of damaged proteins activates stress responses, such as the unfolded protein response (UPRmt), which, while protective, diverts energy away from ATP production. Moreover, oxidized proteins can promote mitochondrial permeability transition pore (mPTP) opening, causing mitochondrial swelling and depolarization, which severely impairs ATP synthesis and can lead to cell death.
In summary, while proteins can fuel cellular respiration, their oxidation has a profound negative impact on ATP generation in mitochondria. Oxidative modification of ETC complexes, citric acid cycle enzymes, and other mitochondrial proteins disrupts energy production pathways, reduces efficiency, and triggers stress responses that further compromise ATP synthesis. Understanding these mechanisms is crucial for addressing metabolic disorders and conditions where protein oxidation is elevated, such as aging, diabetes, and neurodegenerative diseases. Strategies to mitigate protein oxidation, such as enhancing antioxidant defenses or improving protein repair mechanisms, may help preserve mitochondrial function and maintain cellular energy homeostasis.
Where to Buy Warp Fuel in No Man's Sky: A Guide
You may want to see also
Explore related products






![]()
Comparison of protein efficiency versus carbohydrates and fats in respiration
Proteins can indeed be used to fuel cellular respiration, but their efficiency in this process is significantly lower compared to carbohydrates and fats. Carbohydrates are the body's preferred energy source due to their rapid conversion into glucose, which can be quickly metabolized in the presence of oxygen to produce ATP, the cellular energy currency. This process is highly efficient, yielding approximately 36-38 ATP molecules per glucose molecule. In contrast, proteins are primarily used for structural and enzymatic functions, and their breakdown for energy is a secondary role. When proteins are used for respiration, they must first be deaminated to remove nitrogen, which is then converted into glucose through gluconeogenesis. This multi-step process is less efficient, producing only about 32 ATP molecules per molecule of glucose derived from protein, and it places additional stress on the liver and kidneys.
Fats, on the other hand, are the most energy-dense macronutrient, providing approximately 9 kcal per gram compared to 4 kcal per gram for carbohydrates and proteins. During cellular respiration, fats yield a higher number of ATP molecules per gram, with up to 146 ATP molecules produced per fatty acid molecule. This high energy yield makes fats an efficient fuel source, particularly during prolonged, low-intensity activities. However, the oxidation of fats is a slower process compared to carbohydrates, requiring more oxygen and taking longer to mobilize. This makes fats less suitable for high-intensity, short-duration activities where rapid energy production is essential.
When comparing the efficiency of proteins to fats, proteins are clearly the least efficient fuel source for cellular respiration. The energy cost of breaking down proteins, including the energy required for deamination and gluconeogenesis, reduces their net ATP yield. Additionally, the use of proteins for energy can lead to muscle wasting, as amino acids are diverted from their primary role in muscle repair and growth. Fats, while slower to metabolize, provide a more sustained and energy-dense fuel source without compromising structural proteins.
Carbohydrates offer a balance between efficiency and speed, making them the most effective fuel for high-intensity activities. Their rapid breakdown and high ATP yield ensure that energy demands are met quickly, particularly during anaerobic conditions. Proteins, despite their ability to be used for energy, are inefficient and should not be relied upon as a primary fuel source. Instead, they are better utilized for their essential roles in tissue repair, enzyme function, and hormone production.
In summary, while proteins can technically fuel cellular respiration, their efficiency pales in comparison to carbohydrates and fats. Carbohydrates provide a quick and efficient energy source, fats offer high energy density and sustained fuel, and proteins are best reserved for their structural and functional roles. Understanding these differences is crucial for optimizing energy metabolism and maintaining overall physiological health. For individuals engaging in physical activities, a balanced intake of these macronutrients ensures that the body has the appropriate fuel for varying energy demands.
Where to Find Mountain Dew Game Fuel: Ultimate Guide for Gamers
You may want to see also
Frequently asked questions
Yes, proteins can be used to fuel cellular respiration, but they are not the preferred energy source. Cells primarily use carbohydrates and fats for energy, turning to proteins only when these sources are depleted.
Proteins are broken down into amino acids through digestion or cellular processes. The amino acids are then deaminated to remove nitrogen, and the remaining carbon skeletons can enter metabolic pathways like the citric acid cycle (Krebs cycle) to produce ATP.
No, using proteins for cellular respiration is less efficient than using carbohydrates or fats. Proteins yield fewer ATP molecules per gram, and the process produces nitrogenous waste that must be eliminated by the body.
Cells rely on proteins for cellular respiration during prolonged starvation, intense exercise, or when carbohydrate and fat stores are insufficient. This is a last resort mechanism to maintain energy production.
Using proteins for energy can lead to muscle wasting, as structural proteins are broken down, and increased strain on the kidneys due to the need to excrete nitrogenous waste products like urea.































