Vesicle transportation, a fundamental process in cellular biology, relies on a complex interplay of energy sources and molecular mechanisms to ensure the precise movement of cargo within and between cells. At the heart of this process lies adenosine triphosphate (ATP), the primary energy currency of cells, which powers the motor proteins responsible for vesicle trafficking along the cytoskeleton. Additionally, GTP (guanosine triphosphate) plays a critical role by activating small GTPases, such as Rab and Arf proteins, which regulate vesicle budding, docking, and fusion. The hydrolysis of these nucleotide triphosphates provides the necessary energy for conformational changes in proteins, enabling the dynamic assembly and disassembly of vesicle coats and SNARE complexes. Together, these energy-driven mechanisms ensure the efficient and targeted delivery of vesicles, facilitating essential cellular functions like signal transduction, membrane remodeling, and nutrient transport.
| Characteristics | Values |
|---|---|
| Energy Source | ATP (Adenosine Triphosphate) |
| Primary Mechanism | Hydrolysis of ATP by ATPases (e.g., proton pumps, ABC transporters) |
| Key Proteins | SNARE proteins (e.g., VAMP, Syntaxin, SNAP-25), Dynamin, Rab GTPases |
| Types of Transport | Exocytosis (vesicle fusion with plasma membrane), Endocytosis (vesicle formation from plasma membrane), Intracellular trafficking (e.g., Golgi to ER) |
| Driving Force | Electrochemical gradients (e.g., H+, Ca2+), Membrane potential |
| Regulation | Calcium ions (Ca2+) for exocytosis, GTP hydrolysis by Rab proteins, Phosphorylation/dephosphorylation of proteins |
| Organelles Involved | Endoplasmic Reticulum (ER), Golgi apparatus, Lysosomes, Endosomes, Plasma membrane |
| Function | Transport of molecules (e.g., proteins, lipids, neurotransmitters), Cell signaling, Membrane remodeling |
| Disorders Related to Defects | Neurodegenerative diseases (e.g., Alzheimer's, Parkinson's), Lysosomal storage disorders, Diabetes (insulin vesicle release) |
| Recent Advances | Role of lipid composition in vesicle budding, Mechanisms of vesicle tethering and docking, Cryo-EM structures of transport complexes |
Explore related products






What You'll Learn
- ATP Hydrolysis: Energy from ATP powers motor proteins for vesicle movement along cytoskeleton
- GTPases: GTP-binding proteins regulate vesicle budding, docking, and fusion processes
- Membrane Potential: Electrochemical gradients drive ion pumps, indirectly supporting vesicle transport
- Motor Proteins: Kinesin, dynein, and myosin transport vesicles along microtubules and actin
- Coat Proteins: Clathrin and COPI coats shape vesicles and mediate cargo selection
![]()
ATP Hydrolysis: Energy from ATP powers motor proteins for vesicle movement along cytoskeleton
ATP hydrolysis is the cellular process that converts ATP (adenosine triphosphate) into ADP (adenosine diphosphate) and inorganic phosphate, releasing energy in the process. This energy is essential for powering motor proteins, such as kinesins and dyneins, which are responsible for vesicle movement along the cytoskeleton. Imagine a molecular railway system where vesicles, carrying vital cargo like proteins and neurotransmitters, are transported to their precise destinations within the cell. ATP acts as the fuel for this system, enabling motor proteins to "walk" along microtubules, the tracks of the cytoskeleton. Without ATP, this intricate transport mechanism would grind to a halt, disrupting cellular functions like signal transmission and enzyme delivery.
To understand the mechanics, consider the step-by-step process: ATP binds to the motor protein, causing a conformational change that allows the protein to attach to a microtubule. Hydrolysis of ATP to ADP and phosphate then provides the energy for the protein to move along the microtubule, pulling the vesicle with it. For example, kinesin proteins typically move vesicles toward the cell periphery (plus-end directed), while dyneins transport them toward the cell center (minus-end directed). This directional specificity ensures that vesicles reach their correct targets, such as the plasma membrane for exocytosis or the Golgi apparatus for further processing. The efficiency of this system relies on the continuous availability of ATP, which is regenerated through cellular respiration and other metabolic pathways.
From a practical standpoint, disruptions in ATP hydrolysis or motor protein function can have severe consequences. For instance, mutations in kinesin or dynein genes are linked to neurological disorders like Charcot-Marie-Tooth disease, where impaired vesicle transport leads to nerve degeneration. Similarly, conditions that deplete cellular ATP, such as ischemia or metabolic stress, can halt vesicle movement, causing cellular dysfunction. Researchers are exploring ways to modulate ATP levels or enhance motor protein activity to treat such disorders. For example, compounds that stabilize microtubules or increase ATP production are being investigated as potential therapies.
Comparatively, ATP hydrolysis in vesicle transport shares similarities with other cellular processes powered by ATP, such as muscle contraction and DNA replication. However, the precision required for vesicle transport is unparalleled, as motor proteins must navigate a complex cytoskeletal network while carrying cargo. This highlights the adaptability of ATP as an energy source, capable of fueling diverse cellular functions with high specificity. By studying ATP hydrolysis in vesicle transport, scientists gain insights into broader principles of cellular energetics and motility, which could inform advancements in biotechnology and medicine.
In conclusion, ATP hydrolysis is the cornerstone of vesicle transport, providing the energy needed for motor proteins to move vesicles along the cytoskeleton. This process is not only fundamental to cellular function but also a target for therapeutic intervention in diseases caused by transport defects. Understanding the mechanics and implications of ATP-powered vesicle movement offers a window into the elegance and complexity of cellular machinery, underscoring the importance of energy management in biological systems.
Does Volt Need Premium Fuel? Unraveling the Truth for Efficiency
You may want to see also
Explore related products






![]()
GTPases: GTP-binding proteins regulate vesicle budding, docking, and fusion processes
Vesicle transport, the cellular process responsible for moving molecules within and between cells, relies heavily on the precise regulation of budding, docking, and fusion events. At the heart of this regulation are GTPases, a family of proteins that bind and hydrolyze guanosine triphosphate (GTP). These proteins act as molecular switches, cycling between active (GTP-bound) and inactive (GDP-bound) states to control the dynamic assembly and disassembly of protein complexes required for vesicle trafficking.
Consider the Rab GTPases, a subfamily critical for vesicle docking and fusion. When a vesicle approaches its target membrane, Rab proteins on the vesicle surface are activated by GTP binding. This activation recruits effector proteins, such as tethering factors, which position the vesicle correctly for fusion. For instance, Rab5 is essential for early endosome fusion, while Rab7 regulates trafficking to late endosomes. Inhibition of Rab GTPases, often achieved experimentally using dominant-negative mutants or GTPase-activating proteins (GAPs), disrupts these processes, highlighting their indispensable role. Practical applications of this knowledge include targeted therapies for diseases like cancer, where disrupting vesicle trafficking in tumor cells can inhibit growth.
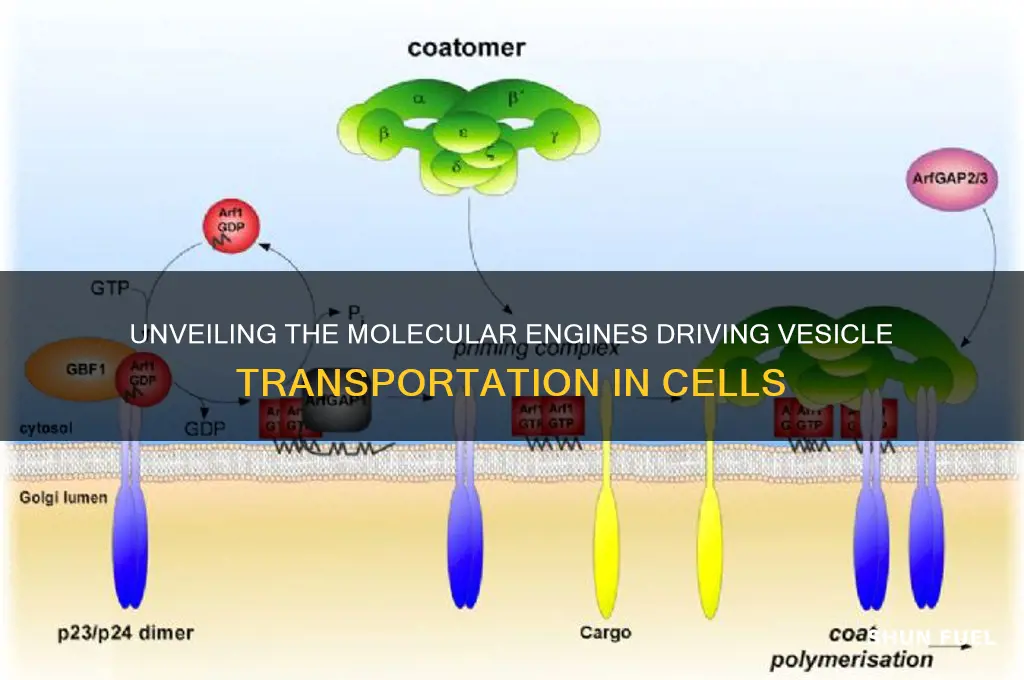
Another key player is the ARF (ADP-ribosylation factor) family of GTPases, which primarily regulate vesicle budding. ARF proteins cycle between the cytosol and membranes, where they recruit coat proteins like COPI and clathrin to deform the membrane and initiate budding. For example, ARF1 is crucial for the formation of COPI-coated vesicles in Golgi-to-ER transport. Activation of ARF GTPases is tightly controlled by guanine nucleotide exchange factors (GEFs) and GAPs, ensuring that budding occurs only when and where needed. Researchers have exploited this mechanism by designing small-molecule inhibitors of ARF GEFs, which show promise in blocking viral replication by disrupting vesicle-mediated entry pathways.
The dynamin family of GTPases provides a striking example of how GTP hydrolysis directly drives mechanical work. Dynamin forms helical structures around the necks of budding vesicles, and its GTPase activity powers the constriction and fission of the membrane. This process is particularly critical in clathrin-mediated endocytosis, where dynamin severs the vesicle from the plasma membrane. Mutations in dynamin genes, such as those causing Charcot-Marie-Tooth disease, underscore the clinical relevance of these proteins. To study dynamin function, researchers often use GTP analogs like GMP-PCP, which stabilize the protein in its active state, allowing for detailed structural and functional analysis.
In summary, GTPases are the molecular orchestrators of vesicle transport, governing budding, docking, and fusion through their GTP-dependent activity. Their role as switches, coupled with the specificity of their effectors, ensures the fidelity and efficiency of intracellular trafficking. Understanding these mechanisms not only advances basic cell biology but also opens avenues for therapeutic intervention in diseases where vesicle transport is dysregulated. Whether through targeted inhibitors, genetic modulation, or structural studies, GTPases remain a focal point in the quest to control the cellular highways of vesicle trafficking.
Deregulation's Role in Harvey's Devastation: A Policy Analysis
You may want to see also
Explore related products





![]()
Membrane Potential: Electrochemical gradients drive ion pumps, indirectly supporting vesicle transport
The movement of vesicles within cells is a complex process, and one of the key drivers is the electrochemical gradient established by membrane potential. This gradient is created by the uneven distribution of ions across the cell membrane, with a higher concentration of positively charged ions outside the cell and a higher concentration of negatively charged ions inside. The resulting voltage difference, typically around -60 to -80 millivolts, provides the energy required for various cellular processes, including vesicle transport.
The Role of Ion Pumps
At the heart of this process are ion pumps, such as the Na+/K+ ATPase pump, which actively transport ions against their concentration gradients. This pump extrudes 3 Na+ ions from the cell while importing 2 K+ ions, consuming 1 ATP molecule in the process. The resulting electrochemical gradient not only maintains the membrane potential but also provides a source of energy for secondary active transporters, like the Na+/Ca2+ exchanger and the Na+/glucose cotransporter. These transporters harness the energy from the Na+ gradient to move other molecules, including those involved in vesicle formation and trafficking.
Indirect Support for Vesicle Transport
While ion pumps do not directly interact with vesicles, their activity is crucial for creating the conditions necessary for vesicle transport. For instance, the calcium ions (Ca2+) regulated by the Na+/Ca2+ exchanger play a critical role in vesicle fusion and release. A rapid increase in intracellular Ca2+ concentration, from approximately 100 nM to 1 μM, triggers the binding of synaptotagmin to phospholipids, facilitating vesicle docking and fusion. This process, known as calcium-triggered exocytosis, is essential for neurotransmitter release at synapses and hormone secretion in endocrine cells.
Practical Implications and Considerations
Understanding the relationship between membrane potential and vesicle transport has significant implications for medical research and therapeutic development. For example, drugs targeting ion pumps or channels, such as diuretics (e.g., thiazides) and anti-epileptics (e.g., carbamazepine), can indirectly affect vesicle transport by altering the electrochemical gradient. Clinicians must consider these effects when prescribing medications, particularly in patients with neurological or endocrine disorders. Moreover, researchers can exploit this knowledge to design more effective therapies, such as calcium channel blockers for treating hypertension or Parkinson's disease.
Optimizing Vesicle Transport in Specific Contexts
In certain scenarios, such as in vitro fertilization or gene therapy, optimizing vesicle transport is crucial for success. For instance, during intracytoplasmic sperm injection (ICSI), the precise control of intracellular Ca2+ concentration is essential for activating the egg and initiating embryonic development. Similarly, in gene therapy, efficient vesicle-mediated delivery of nucleic acids requires a thorough understanding of the underlying electrochemical gradients. By manipulating these gradients, researchers can enhance the efficacy of vesicle-based delivery systems, improving outcomes in various biomedical applications. To achieve this, techniques such as electrophysiological recordings and calcium imaging can be employed to monitor and modulate membrane potential, ensuring optimal conditions for vesicle transport.
Understanding Fuel Surcharge Front Loading: Impact and Strategies for Businesses
You may want to see also
Explore related products






![]()
Motor Proteins: Kinesin, dynein, and myosin transport vesicles along microtubules and actin
Vesicle transport within cells is a finely orchestrated process, powered by motor proteins that move cargo along specific tracks. Among these, kinesin, dynein, and myosin are the key players, each with distinct roles and mechanisms. Kinesin and dynein operate on microtubules, while myosin functions on actin filaments, collectively ensuring the precise delivery of vesicles to their cellular destinations.
Mechanisms and Directionality
Kinesin and dynein are microtubule-associated motors with opposing functions: kinesin moves vesicles toward the cell periphery (plus-end directed), while dynein transports them toward the cell center (minus-end directed). This bidirectional system allows for both anterograde and retrograde transport, essential for maintaining cellular homeostasis. Myosin, on the other hand, operates on actin filaments, facilitating shorter-range movements, often in the cell cortex or during endocytosis. Each motor protein binds ATP, hydrolyzing it to generate the mechanical force required for movement, with kinesin consuming approximately 1 ATP molecule per 8-nanometer step along the microtubule.
Practical Implications and Regulation
Understanding these motors is crucial for therapeutic applications, as defects in their function can lead to neurodegenerative diseases like Alzheimer’s or Huntington’s. For instance, kinesin inhibitors are being explored to disrupt vesicle trafficking in cancer cells, starving them of essential nutrients. Conversely, enhancing dynein activity could improve axonal transport in neuronal disorders. Researchers often use in vitro assays, such as optical trapping or fluorescence microscopy, to study motor protein dynamics, with experiments typically conducted at 37°C to mimic physiological conditions and ATP concentrations of 1–5 mM to ensure optimal motor activity.
Comparative Efficiency and Load Capacity
While kinesin and dynein share the microtubule track, their efficiencies differ. Kinesin moves at speeds of 1–2 micrometers per second, whereas dynein is slower but can carry heavier cargo, up to 10 times the load of kinesin. Myosin, operating on actin, is versatile, with over 35 isoforms tailored to specific cellular tasks, such as muscle contraction or vesicle tethering. This diversity highlights the adaptability of motor proteins to meet varying cellular demands, from rapid signaling to structural support.
Takeaway for Researchers and Clinicians
For those studying vesicle transport, focusing on motor protein interactions with their tracks and cargo provides actionable insights. Techniques like CRISPR-Cas9 can be employed to manipulate motor protein expression in cell lines, while small-molecule modulators offer potential therapeutic avenues. Clinically, understanding these motors’ roles in diseases like amyotrophic lateral sclerosis (ALS), where dynein dysfunction is implicated, could lead to targeted interventions. By dissecting the unique contributions of kinesin, dynein, and myosin, researchers can unlock new strategies for diagnosing and treating disorders rooted in vesicle trafficking defects.
Maximize Your Savings: Understanding Fuel Rewards Gallon Limits
You may want to see also
Explore related products






![]()
Coat Proteins: Clathrin and COPI coats shape vesicles and mediate cargo selection
Vesicle transport, the cellular process of moving molecules within and between cells, relies heavily on coat proteins like clathrin and COPI. These proteins act as molecular scaffolds, shaping vesicles and ensuring the precise selection of cargo for transport. Imagine them as the architects and bouncers of the cellular shipping system, dictating what gets packaged and where it goes.
Clathrin, with its distinctive triskelion structure, forms a polyhedral lattice around budding vesicles, primarily at the plasma membrane and trans-Golgi network. This cage-like structure provides the necessary curvature for vesicle formation and acts as a platform for recruiting adaptor proteins. These adaptors, like AP-2, recognize specific sorting signals on cargo molecules, ensuring only the right "packages" are loaded. For instance, clathrin-coated vesicles are crucial for endocytosis, bringing nutrients and signaling molecules into the cell, and for trafficking proteins from the trans-Golgi network to endosomes.
COPI coats, on the other hand, specialize in retrograde transport, moving cargo from the Golgi apparatus back to the endoplasmic reticulum (ER). Unlike clathrin's cage, COPI forms a more flexible, coat-like structure. It recognizes different sorting signals, primarily the KDEL motif found on proteins destined for the ER. This selective recognition ensures that proteins mistakenly transported to the Golgi are efficiently retrieved, maintaining cellular homeostasis.
Understanding the distinct roles of clathrin and COPI highlights the sophistication of vesicle trafficking. Their ability to shape vesicles and mediate cargo selection is fundamental to cellular function, influencing processes from nutrient uptake to protein secretion. Research into these coat proteins not only deepens our understanding of cellular logistics but also holds promise for developing therapeutic strategies targeting diseases associated with vesicle trafficking defects.
Do Trains Run on Fuel? Exploring Locomotive Power Sources
You may want to see also
Frequently asked questions
The primary source of energy for vesicle transportation is adenosine triphosphate (ATP), which powers motor proteins like kinesin and dynein to move vesicles along microtubules.
Motor proteins bind to vesicles and use ATP hydrolysis to generate mechanical force, allowing them to "walk" along cytoskeletal tracks (microtubules or actin filaments) and transport vesicles to their destinations.
The cytoskeleton, composed of microtubules, actin filaments, and intermediate filaments, provides the structural framework and tracks along which motor proteins move vesicles to specific cellular locations.
Yes, there are two main types: active transport (fueled by ATP and motor proteins) and passive transport (driven by diffusion or membrane fusion, often facilitated by GTP-binding proteins like Rab GTPases).
GTP (guanosine triphosphate) is used by proteins like dynamin and Rab GTPases to regulate vesicle budding, fusion, and trafficking, providing energy for membrane remodeling and targeting.































