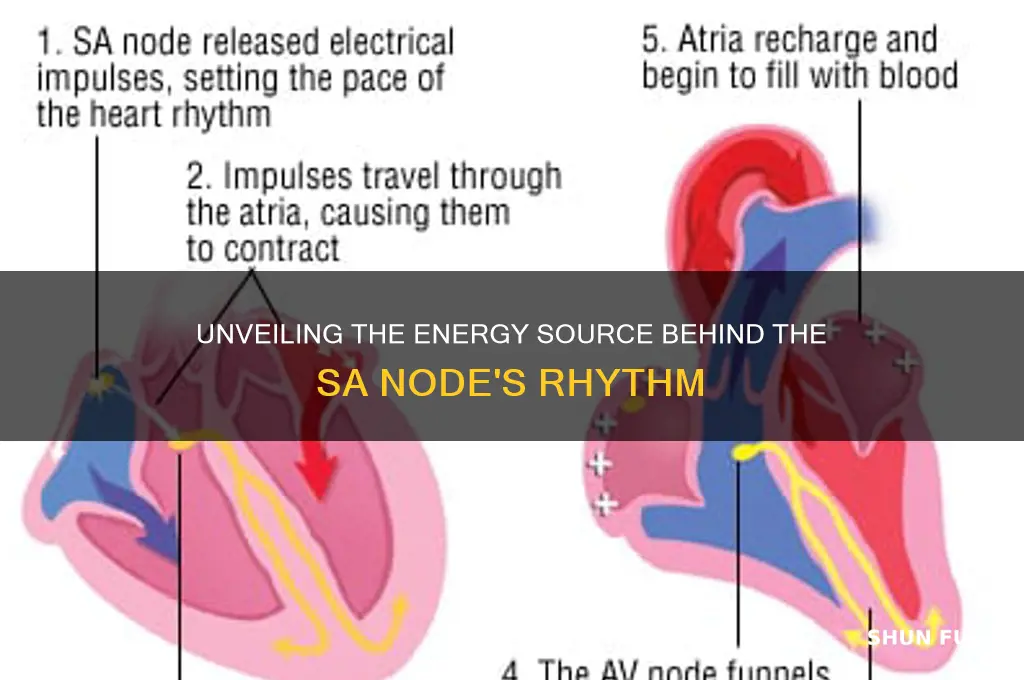
The sinoatrial (SA) node, often referred to as the heart's natural pacemaker, is fueled primarily by a complex interplay of ionic currents and metabolic processes. Its primary energy source is adenosine triphosphate (ATP), which is generated through oxidative phosphorylation in the mitochondria, relying heavily on fatty acid and glucose metabolism. Additionally, the SA node is richly innervated by the autonomic nervous system, with sympathetic stimulation increasing its firing rate via beta-adrenergic receptors and parasympathetic input slowing it through muscarinic receptors. This intricate balance of metabolic and neurohumoral factors ensures the SA node's ability to maintain the heart's rhythmic electrical activity, driving the cardiac cycle efficiently.
| Characteristics | Values |
|---|---|
| Primary Energy Source | ATP (Adenosine Triphosphate) |
| Metabolic Pathways | Primarily oxidative phosphorylation (fatty acid and glucose metabolism) |
| Oxygen Dependence | Highly dependent on aerobic metabolism |
| Fatty Acid Utilization | Preferred substrate under normal conditions |
| Glucose Utilization | Secondary substrate, increases during hypoxia or high workload |
| Lactate Utilization | Minimal, not a significant energy source |
| Mitochondrial Density | High, reflecting reliance on oxidative metabolism |
| Glycogen Stores | Limited, insufficient for prolonged function without oxidative fuel |
| Sensitivity to Ischemia | Highly sensitive; function declines rapidly without oxygen |
| Role of Creatine Phosphate | Provides short-term ATP buffering during transient energy demands |
| Influence of Hormones | Insulin and glucagon modulate substrate selection (e.g., glucose vs. fatty acids) |
| Temperature Sensitivity | Metabolic rate and ATP production increase with temperature |
| pH Dependence | Optimal function at physiological pH (7.35–7.45); acidosis impairs function |
| Calcium Handling | ATP-dependent calcium pumps are critical for pacemaker activity |
| Impact of Hypoxia | Rapidly reduces ATP production, leading to bradycardia or asystole |
| Role of KATP Channels | Activated by low ATP, contributing to rate adaptation during ischemia |
Explore related products






What You'll Learn
- Calcium Ion Flux: Calcium influx via T-type channels triggers depolarization, initiating the action potential
- Sinoatrial Pacemaker Cells: Specialized cells with automaticity drive rhythmic electrical impulses in the SA node
- Potassium Channel Role: Gradual potassium efflux sustains diastolic depolarization, preparing for the next impulse
- Autonomic Nervous Influence: Sympathetic and parasympathetic nerves modulate SA node firing rate
- Intrinsic Automaticity: The SA node’s inherent ability to generate electrical signals without external stimuli
![]()
Calcium Ion Flux: Calcium influx via T-type channels triggers depolarization, initiating the action potential
The sinoatrial (SA) node, often referred to as the heart's natural pacemaker, relies on a delicate interplay of ionic currents to generate electrical impulses. Among these, calcium ion flux plays a pivotal role in initiating the action potential. Specifically, the influx of calcium ions through T-type calcium channels marks the beginning of depolarization, setting the stage for the heart’s rhythmic contractions. This process is not merely a passive event but a finely tuned mechanism that ensures the heart beats consistently and efficiently.
To understand the significance of calcium influx via T-type channels, consider the unique properties of these channels. T-type calcium channels are low-voltage activated, meaning they open at relatively hyperpolarized membrane potentials, typically around -60 mV. This early activation allows calcium ions to enter the cell during the late phase of repolarization, creating a small, inward current known as the pacemaker current (*I*ca-T). This current gradually depolarizes the membrane, eventually reaching the threshold for the opening of L-type calcium channels and sodium channels, which fully depolarize the cell. Without the initial trigger from T-type channels, the SA node’s ability to initiate action potentials would be severely compromised.
From a practical standpoint, understanding this mechanism has implications for medical interventions. For instance, drugs that modulate T-type calcium channels, such as mibefradil (a T-type calcium channel blocker), can alter the heart rate by inhibiting the pacemaker current. While mibefradil is no longer widely used due to side effects, newer compounds targeting T-type channels are being explored for treating arrhythmias. For patients with conditions like sinus node dysfunction, where the SA node fails to maintain a normal heart rate, therapies aimed at enhancing or restoring calcium influx could be transformative. However, caution must be exercised, as excessive blockade of T-type channels can lead to bradycardia, highlighting the need for precise dosing and monitoring.
Comparatively, the role of calcium influx in the SA node contrasts with its function in other cardiac cells, such as ventricular myocytes. In ventricular cells, calcium influx primarily occurs through L-type calcium channels during the plateau phase of the action potential, facilitating contraction. In the SA node, however, T-type channels take center stage during the early phases of depolarization, emphasizing the node’s specialized role in rhythm generation. This distinction underscores the importance of calcium ion flux as a key differentiator in cardiac electrophysiology.
In conclusion, calcium influx via T-type channels is not just a component of SA node function—it is the spark that ignites the heart’s electrical activity. By triggering depolarization, this mechanism ensures the heart’s rhythmic beating, making it a critical target for both physiological understanding and therapeutic intervention. Whether in the context of drug development or clinical management, appreciating the nuances of calcium ion flux in the SA node is essential for advancing cardiac care.
Understanding Hybrid Vehicles: Exploring the Fuel Types That Power Them
You may want to see also
Explore related products






![]()
Sinoatrial Pacemaker Cells: Specialized cells with automaticity drive rhythmic electrical impulses in the SA node
The sinoatrial (SA) node, often referred to as the heart's natural pacemaker, relies on specialized cells known as sinoatrial pacemaker cells to initiate and maintain its rhythmic electrical impulses. These cells possess a unique property called automaticity, which allows them to spontaneously depolarize and generate action potentials without external stimulation. This intrinsic ability is what fuels the SA node, ensuring the heart beats consistently and efficiently. Unlike other cardiac cells, pacemaker cells have a distinct ion channel composition, particularly a higher density of hyperpolarization-activated cyclic nucleotide-gated (HCN) channels, which are critical for their automaticity.
To understand how these cells fuel the SA node, consider their electrophysiological behavior. During the diastolic phase, pacemaker cells gradually depolarize due to the influx of sodium and calcium ions, primarily through HCN channels (also known as "funny" current, If). This slow depolarization continues until the threshold potential is reached, triggering a rapid depolarization phase driven by calcium influx. The action potential then propagates through the heart, initiating contraction. Notably, the resting membrane potential of pacemaker cells is less negative than that of other cardiac cells, typically around -60 mV, which facilitates their automaticity. This process repeats approximately 60–100 times per minute in a healthy adult, depending on physiological demands.
From a practical perspective, understanding the fuel for the SA node is crucial in clinical settings, particularly when addressing arrhythmias or heart block. For instance, drugs like beta-blockers or calcium channel blockers can modulate the automaticity of pacemaker cells by altering ion channel activity. Beta-blockers reduce the heart rate by decreasing the inward current through HCN channels, while calcium channel blockers slow conduction by reducing calcium influx. Conversely, in cases of bradycardia, pacing devices or drugs like isoproterenol (a beta-adrenergic agonist) can enhance the automaticity of pacemaker cells, increasing heart rate. These interventions highlight the importance of targeting the unique properties of SA node cells to manage cardiac rhythm effectively.
Comparatively, the automaticity of SA node cells contrasts with other cardiac tissues, such as atrial or ventricular myocytes, which rely on external electrical signals for activation. This distinction underscores the SA node's role as the primary pacemaker. However, if the SA node fails, subsidiary pacemakers in the atrioventricular (AV) node or His-Purkinje system can take over, albeit at a slower rate. This hierarchical system ensures cardiac continuity but emphasizes the SA node's dominance due to its faster intrinsic rate. Thus, the specialized automaticity of sinoatrial pacemaker cells is not just a feature but a necessity for maintaining optimal cardiac function.
In summary, sinoatrial pacemaker cells fuel the SA node through their inherent automaticity, driven by a unique ion channel profile and electrophysiological behavior. This mechanism ensures the heart's rhythmic electrical impulses, adapting to physiological demands. Clinicians and researchers can leverage this knowledge to develop targeted therapies for arrhythmias, emphasizing the SA node's central role in cardiac health. By focusing on these specialized cells, we gain insights into both the normal functioning and potential dysfunctions of the heart's electrical system.
Unraveling the Roots: What Fuels Conspiracy Theories and Why They Persist
You may want to see also
Explore related products






![]()
Potassium Channel Role: Gradual potassium efflux sustains diastolic depolarization, preparing for the next impulse
The sinoatrial (SA) node, often referred to as the heart's natural pacemaker, relies on a delicate interplay of ionic currents to generate and propagate electrical impulses. Among these, the role of potassium channels is particularly crucial during the diastolic phase. Gradual potassium efflux through these channels sustains diastolic depolarization, a process that quietly prepares the SA node for the next impulse. This slow, controlled release of potassium ions creates a subtle shift in membrane potential, setting the stage for the subsequent action potential. Without this mechanism, the rhythmicity of the heart would falter, underscoring the potassium channel's indispensable role in cardiac function.
To understand this process, consider the SA node's membrane potential during diastole. At rest, the membrane potential hovers around -55 to -60 mV. As potassium ions gradually leak out through potassium channels, the membrane potential slowly rises toward the threshold for activation, typically around -40 mV. This phase, known as diastolic depolarization, is not abrupt but rather a gradual ascent, akin to a climber steadily approaching a summit. The If current (funny current), carried by hyperpolarization-activated cyclic nucleotide-gated (HCN) channels, also contributes, but the potassium efflux is the silent architect, ensuring the membrane potential remains on course. This interplay highlights the precision required for cardiac pacing.
Clinically, disruptions in potassium channel function can have profound implications. For instance, conditions like hyperkalemia (elevated serum potassium) or hypokalemia (low serum potassium) can alter the rate of potassium efflux, thereby affecting diastolic depolarization. In hyperkalemia, excessive potassium outside the cell can slow the efflux, delaying depolarization and potentially causing bradycardia. Conversely, hypokalemia accelerates efflux, leading to rapid depolarization and possible arrhythmias. Patients on diuretics, particularly loop diuretics, are at risk of hypokalemia and should monitor serum potassium levels regularly, aiming to maintain values between 3.5 and 5.0 mmol/L. These examples illustrate the potassium channel's central role in maintaining cardiac rhythm.
From a pharmacological perspective, drugs targeting potassium channels can modulate SA node activity. For example, potassium channel blockers like 4-aminopyridine can inhibit potassium efflux, slowing diastolic depolarization and reducing heart rate. Conversely, potassium channel openers, such as pinacidil, enhance efflux, which can be beneficial in conditions like hypertension by promoting vasodilation. However, such interventions must be approached cautiously, as altering potassium dynamics can have systemic effects. For instance, in elderly patients (aged 65 and above), drug metabolism may be slower, necessitating lower dosages to avoid adverse effects. This underscores the need for personalized treatment strategies when manipulating potassium channel activity.
In practical terms, understanding the potassium channel's role in diastolic depolarization offers actionable insights for healthcare providers. For patients with arrhythmias, monitoring electrolyte balance, particularly potassium, is critical. Dietary adjustments, such as increasing potassium-rich foods (e.g., bananas, spinach) in hypokalemia or restricting them in hyperkalemia, can complement pharmacotherapy. Additionally, educating patients about the signs of arrhythmias—such as palpitations, dizziness, or chest discomfort—empowers them to seek timely medical intervention. By focusing on the gradual potassium efflux that sustains diastolic depolarization, clinicians can fine-tune therapies to optimize cardiac rhythm and overall heart health.
Mastering Carburetor Fuel Spray: Tips for Optimal Engine Performance
You may want to see also
Explore related products






![]()
Autonomic Nervous Influence: Sympathetic and parasympathetic nerves modulate SA node firing rate
The sinoatrial (SA) node, often called the heart's natural pacemaker, relies on a delicate balance of intrinsic and extrinsic factors to maintain its rhythmic firing. Among these, the autonomic nervous system plays a pivotal role, acting as a dynamic regulator of the SA node's activity. Sympathetic and parasympathetic nerves, the two arms of this system, exert opposing influences on the SA node's firing rate, ensuring the heart adapts to the body's ever-changing demands.
Understanding the Dual Control Mechanism
Sympathetic nerves, originating from the thoracic spinal cord, release norepinephrine (noradrenaline) to increase the SA node's firing rate. This occurs through the activation of beta-1 adrenergic receptors, which enhance the influx of calcium ions and accelerate the depolarization process. For instance, during exercise or stress, sympathetic activation can elevate the heart rate from a resting 60–100 beats per minute (bpm) to 160–200 bpm in healthy adults. Conversely, parasympathetic nerves, via the vagus nerve, release acetylcholine to decrease the firing rate. This is achieved by activating muscarinic receptors, which increase potassium efflux and slow depolarization. At rest, parasympathetic activity can lower the heart rate to around 50–60 bpm in well-conditioned individuals, such as athletes.
Practical Implications and Modulation
Understanding this dual control is crucial for managing heart rate in clinical settings. Beta-blockers, which antagonize beta-1 receptors, are commonly prescribed to reduce sympathetic influence in conditions like hypertension or arrhythmias. For example, a 20–40 mg daily dose of propranolol can effectively lower heart rate by 10–20%. Conversely, in cases of bradycardia (slow heart rate), atropine, an anticholinergic agent, can be administered to block parasympathetic activity, increasing the SA node's firing rate. Dosage typically starts at 0.5–1 mg intravenously, titrated based on response.
Comparative Analysis: Sympathetic vs. Parasympathetic Dominance
The interplay between these systems is particularly evident in scenarios like the "fight or flight" response, where sympathetic dominance prevails, versus relaxation or sleep, where parasympathetic activity dominates. For instance, during a panic attack, excessive sympathetic stimulation can cause heart rates to spike above 180 bpm, while deep breathing exercises activate the parasympathetic system, reducing heart rate by 10–15% within minutes. This highlights the importance of lifestyle interventions, such as mindfulness or yoga, in modulating autonomic balance.
Takeaway: Harnessing Autonomic Influence for Heart Health
To optimize SA node function, individuals can adopt strategies that balance sympathetic and parasympathetic activity. Regular aerobic exercise, for example, enhances parasympathetic tone over time, leading to a lower resting heart rate. Conversely, limiting caffeine and stress management techniques, like progressive muscle relaxation, can mitigate excessive sympathetic activation. For older adults (ages 65+), who often experience reduced parasympathetic activity, gentle exercises like tai chi or walking can help maintain autonomic balance. By understanding and influencing these neural pathways, one can effectively "fuel" the SA node for sustained cardiac health.
Understanding Fuel Points: How They Work and Maximize Savings
You may want to see also
Explore related products






![]()
Intrinsic Automaticity: The SA node’s inherent ability to generate electrical signals without external stimuli
The sinoatrial (SA) node, often referred to as the heart's natural pacemaker, operates with a remarkable degree of autonomy. Unlike other cells in the body that rely on external signals to initiate activity, the SA node possesses intrinsic automaticity—the inherent ability to generate electrical signals spontaneously. This self-starting mechanism is fueled by a complex interplay of ion channels and cellular processes, ensuring the heart beats rhythmically without external intervention. Understanding this intrinsic property is crucial for appreciating the heart's reliability and for diagnosing disorders that disrupt its natural rhythm.
At the core of the SA node's automaticity is the funny current (If), a unique ionic current mediated by hyperpolarization-activated cyclic nucleotide-gated (HCN) channels. These channels allow sodium and potassium ions to flow into the cell during diastolic depolarization, gradually increasing the cell's membrane potential until it reaches the threshold for an action potential. This process is not triggered by external stimuli but by the cell's own internal mechanisms. The If current is particularly sensitive to cyclic adenosine monophosphate (cAMP), a secondary messenger that modulates the channel's activity. For instance, increased sympathetic stimulation elevates cAMP levels, accelerating the depolarization rate and thus the heart rate.
To illustrate the practical implications of intrinsic automaticity, consider the impact of medications like beta-blockers or calcium channel blockers. Beta-blockers reduce cAMP levels by inhibiting beta-adrenergic receptors, slowing the If current and decreasing heart rate. Conversely, calcium channel blockers reduce the inward calcium current during the plateau phase of the action potential, indirectly affecting the SA node's firing rate. These interventions highlight how external agents can modulate, but not initiate, the SA node's intrinsic rhythm. This distinction is vital for clinicians tailoring treatments for arrhythmias or heart rate disorders.
A comparative analysis of the SA node and other cardiac tissues underscores its uniqueness. While atrial and ventricular myocytes rely on external electrical impulses to depolarize, the SA node's automaticity ensures it acts as the heart's primary pacemaker. This autonomy is not absolute, however; the autonomic nervous system can modulate its activity. For example, during exercise, increased sympathetic activity accelerates the SA node's firing rate, while parasympathetic stimulation via the vagus nerve slows it down. Yet, even in the absence of neural input, the SA node continues to generate electrical signals, a testament to its intrinsic nature.
In practical terms, understanding intrinsic automaticity is essential for interpreting electrocardiograms (ECGs) and diagnosing rhythm disorders. A healthy SA node produces a consistent P wave morphology on an ECG, reflecting its regular firing. Deviations from this pattern, such as sinus arrhythmia or sinoatrial block, indicate dysfunction in the node's automaticity. For patients, this knowledge empowers them to recognize symptoms like palpitations or dizziness as potential signs of SA node issues. Clinicians, meanwhile, can use this understanding to prescribe targeted therapies, such as pacemaker implantation for severe sinoatrial node dysfunction.
In conclusion, the SA node's intrinsic automaticity is a marvel of biological engineering, ensuring the heart's rhythmic function without external reliance. By focusing on the mechanisms driving this property, from the If current to autonomic modulation, we gain insights into both normal cardiac physiology and pathological conditions. This knowledge not only advances medical science but also translates into practical applications for patient care, making it a cornerstone of cardiovascular health.
Understanding the Fuel Efficiency of the Do 27 Aircraft
You may want to see also
Frequently asked questions
The primary fuel source for the SA node (sinoatrial node) is adenosine triphosphate (ATP), which is generated primarily through oxidative phosphorylation using fatty acids and glucose.
The SA node relies more on fatty acids for energy under normal conditions, as they are the preferred substrate for oxidative phosphorylation in cardiac cells, including the SA node.
The SA node cannot function without oxygen because it primarily depends on aerobic metabolism (oxidative phosphorylation) to produce ATP. Anaerobic metabolism (glycolysis) is inefficient and cannot sustain the high energy demands of the SA node.