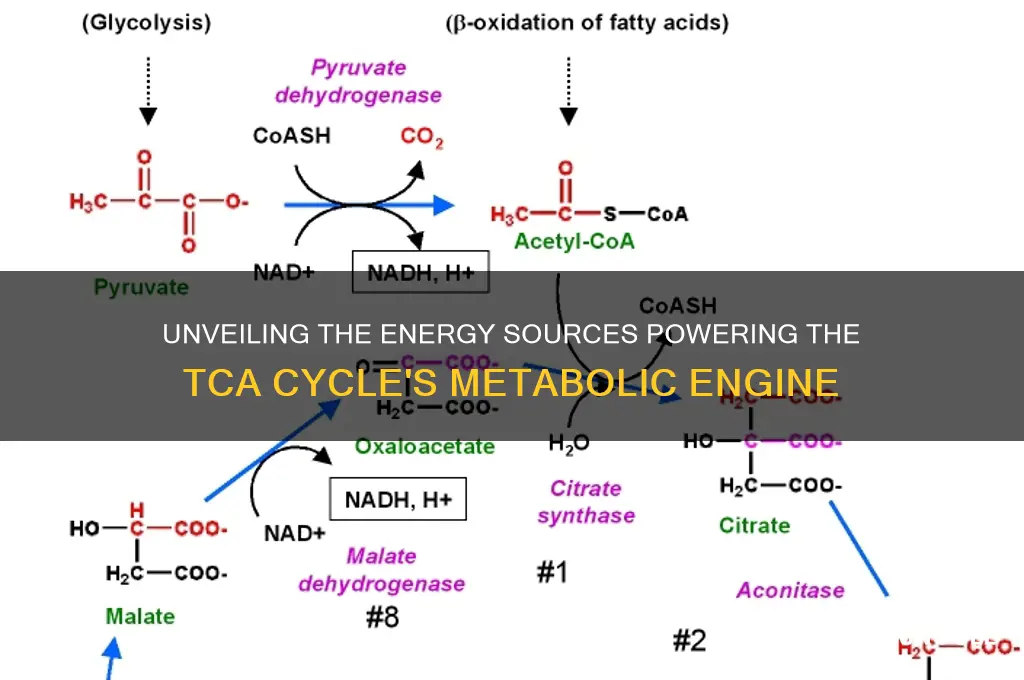
The TCA cycle, also known as the citric acid cycle or Krebs cycle, is a central metabolic pathway that generates energy by oxidizing acetyl-CoA derived from carbohydrates, fats, and proteins. This cycle is fueled primarily by acetyl-CoA, which is produced during the breakdown of glucose (via glycolysis), fatty acids (through beta-oxidation), and amino acids (via various catabolic pathways). Once inside the mitochondria, acetyl-CoA combines with oxaloacetate to form citrate, initiating the cycle. As the cycle progresses, high-energy molecules like NADH and FADH₂ are produced, which ultimately drive ATP synthesis through oxidative phosphorylation. Thus, the TCA cycle serves as a critical hub for energy production, linking the metabolism of diverse macronutrients to cellular respiration.
| Characteristics | Values |
|---|---|
| Primary Fuel | Acetyl-CoA (derived from pyruvate, fatty acids, and amino acids) |
| Carbon Source | Pyruvate (from glucose), fatty acids (from lipids), ketone bodies, and certain amino acids |
| Entry Point | Acetyl-CoA enters the TCA cycle by combining with oxaloacetate to form citrate |
| Key Intermediates | Citrate, isocitrate, α-ketoglutarate, succinyl-CoA, succinate, fumarate, malate, oxaloacetate |
| Energy Carriers | NADH, FADH2, and ATP (or GTP) produced during the cycle |
| Coenzymes Required | NAD+, FAD, and coenzyme A (CoA) |
| Location | Mitochondrial matrix in eukaryotes; cytosol in prokaryotes |
| Oxygen Requirement | Aerobic (requires oxygen for electron transport chain to regenerate NAD+ and FAD) |
| Net ATP Yield per Glucose | ~30-32 ATP (including oxidative phosphorylation) |
| Regulation | Controlled by enzyme activity (e.g., citrate synthase, isocitrate dehydrogenase) and substrate availability |
| Role in Metabolism | Central hub for carbohydrate, fat, and protein metabolism; provides precursors for biosynthesis |
| Alternative Names | Krebs cycle, citric acid cycle |
Explore related products






What You'll Learn
- Pyruvate Oxidation: Converts pyruvate to acetyl-CoA, entering the TCA cycle for energy production
- Fatty Acid Breakdown: Beta-oxidation generates acetyl-CoA from fats, fueling the TCA cycle
- Amino Acid Degradation: Certain amino acids are converted to TCA intermediates, sustaining the cycle
- Glucose Metabolism: Glycolysis produces pyruvate, which feeds into the TCA cycle
- Ketone Bodies: During fasting, ketones provide acetyl-CoA for the TCA cycle
![]()
Pyruvate Oxidation: Converts pyruvate to acetyl-CoA, entering the TCA cycle for energy production
Pyruvate oxidation stands as the critical gateway to the TCA cycle, transforming pyruvate—the end product of glycolysis—into acetyl-CoA, the molecule that fuels this central metabolic pathway. This process occurs in the mitochondrial matrix and involves a series of enzymatic reactions that not only generate acetyl-CoA but also produce NADH, a key electron carrier in oxidative phosphorylation. Without pyruvate oxidation, the TCA cycle would lack its primary substrate, halting energy production in aerobic organisms.
Consider the step-by-step mechanism: pyruvate dehydrogenase complex (PDC) catalyzes the oxidative decarboxylation of pyruvate, stripping off a carbon dioxide molecule and transferring the remaining acetyl group to coenzyme A, forming acetyl-CoA. This reaction is irreversible and tightly regulated, ensuring a unidirectional flow into the TCA cycle. Notably, PDC requires five cofactors—thiamine pyrophosphate (TPP), lipoamide, FAD, NAD+, and CoA—highlighting the metabolic interconnectedness of this process. For instance, a deficiency in thiamine (vitamin B1) can impair PDC activity, leading to energy deficits, particularly in high-energy-demand tissues like the brain and muscles.
From a practical standpoint, optimizing pyruvate oxidation can enhance metabolic efficiency, especially in athletes or individuals with high energy demands. Dietary strategies include consuming adequate B vitamins (B1, B2, B3) to support PDC cofactors, as well as maintaining a balanced intake of carbohydrates to ensure a steady supply of pyruvate. For example, a pre-workout meal rich in complex carbohydrates (e.g., whole grains, fruits) can provide a sustained release of glucose, ultimately fueling pyruvate production and subsequent acetyl-CoA formation. Conversely, in conditions like diabetes or metabolic syndrome, impaired pyruvate oxidation can exacerbate insulin resistance, emphasizing the need for targeted nutritional interventions.
Comparatively, pyruvate oxidation contrasts with anaerobic glycolysis, which bypasses the TCA cycle and produces lactate under oxygen-limited conditions. While glycolysis yields only 2 ATP per glucose molecule, the TCA cycle and oxidative phosphorylation generate up to 36–38 ATP, underscoring the efficiency of aerobic metabolism. This distinction is particularly relevant in endurance sports, where training adaptations increase mitochondrial density and PDC activity, allowing athletes to rely more on pyruvate oxidation and less on glycolysis, thereby delaying fatigue.
In conclusion, pyruvate oxidation is not merely a biochemical step but a metabolic linchpin that bridges glycolysis and the TCA cycle. Its regulation, efficiency, and interplay with dietary factors make it a critical target for optimizing energy production, whether in health, disease, or performance settings. Understanding this process empowers individuals to make informed decisions about nutrition, supplementation, and lifestyle, ensuring that the TCA cycle operates at its full potential.
Ethanol Fuel Emissions: What’s Released and Environmental Impact Explained
You may want to see also
Explore related products



![Essencea Citric Acid 5LB Pure Bulk Ingredients | Non-GMO | 100% Pure Citric Acid Powder [Packaging May Vary]](https://m.media-amazon.com/images/I/51O7Lk96ljL._AC_UL320_.jpg)


![]()
Fatty Acid Breakdown: Beta-oxidation generates acetyl-CoA from fats, fueling the TCA cycle
The human body is a metabolic powerhouse, capable of extracting energy from a variety of sources. One of the most efficient ways it accomplishes this is through the breakdown of fatty acids, a process that ultimately fuels the TCA cycle. Beta-oxidation, a series of enzymatic reactions occurring in the mitochondrial matrix, plays a pivotal role in this process by cleaving fatty acyl-CoA molecules into acetyl-CoA units. Each round of beta-oxidation shortens the fatty acid chain by two carbon atoms, releasing one molecule of acetyl-CoA, NADH, and FADH₂. This acetyl-CoA then enters the TCA cycle, where it is further oxidized to release energy in the form of ATP.
Consider the example of a 16-carbon fatty acid like palmitoyl-CoA. Through seven cycles of beta-oxidation, it yields eight molecules of acetyl-CoA, seven NADH, and seven FADH₂. These acetyl-CoA molecules then enter the TCA cycle, driving the production of approximately 106 ATP molecules per molecule of palmitate. This highlights the remarkable efficiency of fatty acid breakdown as a fuel source for cellular energy production. For individuals engaging in prolonged physical activity or those on low-carbohydrate diets, this pathway becomes particularly crucial, as it provides a sustained and substantial energy supply.
From a practical standpoint, understanding beta-oxidation and its role in fueling the TCA cycle can inform dietary and lifestyle choices. For instance, consuming healthy fats like avocados, nuts, and olive oil ensures a steady supply of fatty acids for beta-oxidation. However, excessive fat intake without adequate physical activity can lead to lipid accumulation, disrupting metabolic balance. Athletes and fitness enthusiasts can optimize their energy levels by timing fat intake around endurance activities, allowing beta-oxidation to meet increased energy demands. Conversely, individuals with metabolic disorders, such as fatty acid oxidation defects, may require specialized diets to manage their condition effectively.
A comparative analysis reveals the advantages of fatty acid breakdown over glucose metabolism. While glucose yields only two molecules of acetyl-CoA per molecule, fatty acids produce multiples of acetyl-CoA depending on their chain length. This makes fats a more energy-dense fuel source, particularly during fasting or low-carbohydrate states. However, beta-oxidation is not without limitations. Unlike glucose, fatty acids cannot be converted into glucose via gluconeogenesis, making them unsuitable for fueling certain tissues like the brain under normal conditions. Additionally, the process requires oxygen, rendering it inefficient in anaerobic environments.
In conclusion, beta-oxidation serves as a critical bridge between fatty acid breakdown and the TCA cycle, ensuring a continuous supply of acetyl-CoA for energy production. By understanding this process, individuals can make informed decisions about their diet and activity levels to optimize metabolic efficiency. Whether for athletic performance, weight management, or metabolic health, harnessing the power of beta-oxidation can unlock the full potential of fats as a fuel source.
Easy Steps to Remove Your Vehicle's Fuel Door Safely
You may want to see also
Explore related products






![]()
Amino Acid Degradation: Certain amino acids are converted to TCA intermediates, sustaining the cycle
Amino acids, the building blocks of proteins, play a dual role in cellular metabolism: they are essential for protein synthesis, but they also serve as a critical fuel source for the tricarboxylic acid (TCA) cycle, particularly during periods of carbohydrate scarcity or increased energy demand. Certain amino acids, such as glutamate, aspartate, and branched-chain amino acids (BCAAs) like leucine, isoleucine, and valine, are uniquely positioned to enter the TCA cycle after degradation. This process not only provides energy but also ensures the cycle’s continuity by replenishing key intermediates like α-ketoglutarate and oxaloacetate. For instance, glutamate is transaminated to form α-ketoglutarate, a direct TCA cycle intermediate, while BCAAs are catabolized to produce acetyl-CoA and succinyl-CoA, further fueling oxidative phosphorylation.
Consider the metabolic demands of athletes or individuals on low-carbohydrate diets. During prolonged exercise or fasting, muscle protein breakdown increases, releasing amino acids into the bloodstream. These amino acids are then degraded in the liver, with their carbon skeletons entering the TCA cycle to sustain ATP production. For example, leucine degradation yields acetyl-CoA, which enters the cycle at the starting point, while isoleucine and valine produce succinyl-CoA, bypassing the initial steps but still contributing to the cycle’s flux. This mechanism is particularly vital in skeletal muscle, where BCAAs account for approximately 35% of essential amino acids and serve as a significant energy source during endurance activities.
However, reliance on amino acid degradation for TCA cycle fueling is not without caution. Excessive protein breakdown, especially in states of chronic energy deficit or certain metabolic disorders, can lead to muscle wasting and impaired protein synthesis. For instance, in uncontrolled diabetes or severe fasting, elevated glucagon levels promote amino acid catabolism, potentially depleting muscle mass. To mitigate this, individuals should aim to consume adequate carbohydrates and proteins, ensuring a balanced macronutrient intake. A practical tip for athletes is to include a BCAA supplement (5–10 grams) during prolonged training sessions to support energy production without accelerating muscle breakdown.
Comparatively, the role of amino acids in TCA cycle fueling contrasts with that of carbohydrates and fats. While glucose and fatty acids are the primary substrates, amino acids act as a metabolic reserve, activated under specific conditions. For example, during a high-intensity workout, carbohydrates are the preferred fuel, but as glycogen stores deplete, amino acids become increasingly important. This hierarchical utilization underscores the adaptability of cellular metabolism. Interestingly, certain amino acids, like glutamine, also play a regulatory role by modulating TCA cycle enzyme activity, further highlighting their multifaceted contribution.
In practical terms, understanding amino acid degradation can inform dietary strategies for various populations. For older adults, who often experience age-related muscle loss (sarcopenia), ensuring sufficient protein intake (1.0–1.2 g/kg/day) can help preserve muscle mass while still allowing for TCA cycle fueling when needed. Similarly, individuals with metabolic disorders, such as maple syrup urine disease (a defect in BCAA degradation), require careful dietary management to prevent toxic accumulation of amino acids while maintaining energy homeostasis. By recognizing the dual role of amino acids, one can optimize metabolic health and energy production across diverse physiological states.
Does Fuel Additive Expire? Shelf Life and Effectiveness Explained
You may want to see also
Explore related products






![]()
Glucose Metabolism: Glycolysis produces pyruvate, which feeds into the TCA cycle
Glucose, the body's primary energy currency, undergoes a complex journey to fuel cellular processes. The first leg of this journey is glycolysis, a series of reactions occurring in the cytoplasm that breaks down one molecule of glucose into two molecules of pyruvate. This process, while generating a modest amount of ATP, is crucial as it sets the stage for the more energy-rich TCA cycle.
Consider glycolysis as the initial investment in a high-yield energy production system. Under aerobic conditions, pyruvate, the end product of glycolysis, is transported into the mitochondria. Here, it undergoes oxidative decarboxylation, converting into acetyl-CoA, the molecule that directly fuels the TCA cycle. This step is pivotal, as it bridges the gap between glucose breakdown and the subsequent energy extraction processes.
The TCA cycle, also known as the Krebs cycle, is a series of enzymatic reactions in the mitochondrial matrix. Acetyl-CoA, derived from pyruvate, combines with oxaloacetate to form citrate, initiating a cycle that generates reducing equivalents (NADH and FADH2) and releases carbon dioxide. These reducing equivalents are then funneled into the electron transport chain, where they drive ATP synthesis through oxidative phosphorylation.
For optimal TCA cycle function, ensure adequate pyruvate supply by maintaining a balanced glucose intake. While the body can derive pyruvate from other sources like amino acids, glucose remains the primary and most efficient fuel. Athletes and individuals with high energy demands may benefit from carbohydrate-rich diets to sustain pyruvate production. However, excessive glucose intake can lead to metabolic imbalances, emphasizing the importance of moderation.
In summary, glycolysis acts as the gateway to the TCA cycle, transforming glucose into pyruvate, which is then converted into acetyl-CoA. This sequential process underscores the interconnectedness of metabolic pathways and highlights the central role of glucose in cellular energy production. By understanding this mechanism, one can appreciate the precision and efficiency of the body's energy management system.
Understanding Bioethanol Fuel: How It Powers Vehicles and Reduces Emissions
You may want to see also
Explore related products






![]()
Ketone Bodies: During fasting, ketones provide acetyl-CoA for the TCA cycle
During fasting, the body shifts from glucose-dependent metabolism to utilizing alternative energy sources, with ketone bodies emerging as a critical fuel for the TCA cycle. When glycogen stores are depleted, typically after 12–16 hours without food, the liver begins to break down fatty acids into ketones: acetoacetate, β-hydroxybutyrate, and acetone. These ketones serve as a lifeline for tissues like the brain and muscles, which cannot directly metabolize fatty acids for energy. The process hinges on the conversion of acetoacetate to acetyl-CoA, a direct substrate for the TCA cycle, ensuring cellular energy production continues even in the absence of carbohydrates.
The mechanism by which ketones fuel the TCA cycle is both elegant and efficient. β-hydroxybutyrate, the most abundant ketone body, is first converted back to acetoacetate in the mitochondria. Acetoacetate is then enzymatically broken down into acetyl-CoA and acetone. This acetyl-CoA enters the TCA cycle, where it is oxidized to produce ATP, NADH, and FADH2, driving oxidative phosphorylation. For individuals fasting or on ketogenic diets, this pathway becomes the primary means of sustaining energy levels, particularly for the brain, which can derive up to 70% of its energy from ketones during prolonged fasting.
Practical considerations for optimizing ketone utilization during fasting include monitoring the duration of the fast, as ketogenesis typically peaks after 2–3 days. Consuming medium-chain triglycerides (MCTs), found in coconut oil or supplements, can accelerate ketone production due to their rapid conversion in the liver. However, excessive MCT intake may cause gastrointestinal discomfort, so starting with 5–10 grams per day and gradually increasing is advisable. Hydration and electrolyte balance are also critical, as ketosis can lead to diuresis and mineral loss, potentially causing fatigue or cramps.
Comparatively, ketone-derived acetyl-CoA offers a cleaner energy source than glucose, producing fewer reactive oxygen species (ROS) during metabolism. This makes ketosis particularly beneficial for individuals with metabolic disorders or those seeking to reduce oxidative stress. However, transitioning to ketosis can be challenging, with symptoms like "keto flu" (headache, fatigue, irritability) occurring during the first week. These symptoms typically resolve as the body adapts, but gradual dietary adjustments and adequate nutrient intake can ease the transition.
In conclusion, ketone bodies are a vital fuel for the TCA cycle during fasting, providing a sustainable energy source when glucose is scarce. Understanding the biochemistry behind ketogenesis and its integration into cellular metabolism empowers individuals to harness this pathway effectively. Whether for weight management, metabolic health, or cognitive enhancement, optimizing ketone utilization requires a balanced approach, combining dietary strategies with awareness of physiological responses. By embracing this metabolic flexibility, one can unlock the full potential of ketones as a powerful energy substrate.
Does Air Conditioning Use Fuel? Exploring AC's Impact on Mileage
You may want to see also
Frequently asked questions
The primary fuel source for the TCA cycle (tricarboxylic acid cycle or Krebs cycle) is acetyl-CoA, which is derived from the breakdown of carbohydrates, fats, and proteins.
Glucose fuels the TCA cycle by first being broken down into pyruvate through glycolysis. Pyruvate is then converted to acetyl-CoA, which enters the TCA cycle as its primary substrate.
Yes, fatty acids and amino acids can fuel the TCA cycle. Fatty acids are broken down through beta-oxidation to produce acetyl-CoA, while certain amino acids are converted into intermediates of the TCA cycle, such as alpha-ketoglutarate or oxaloacetate.