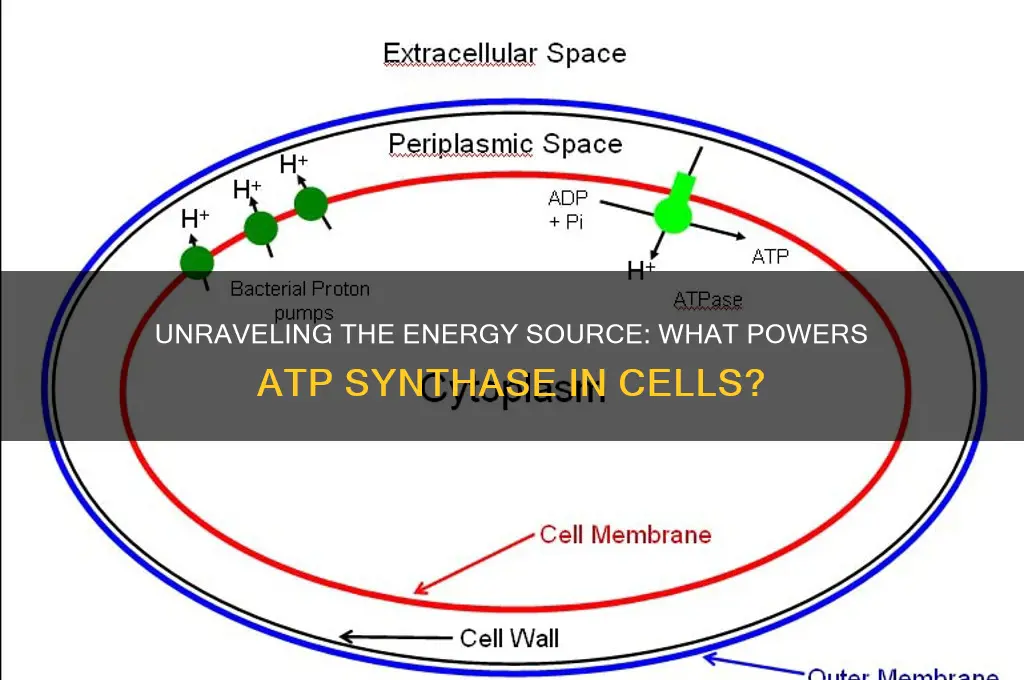
ATP synthase, a crucial enzyme in cellular energy production, is fueled primarily by the proton gradient established across the inner mitochondrial membrane or the thylakoid membrane in chloroplasts. This proton gradient, generated by the electron transport chain during oxidative phosphorylation or photosynthesis, drives the rotation of the enzyme’s rotor subunit, known as the F₀ component. As protons flow back into the matrix or stroma through ATP synthase, their energy is harnessed to phosphorylate ADP to ATP, the cell’s primary energy currency. This process, known as chemiosmosis, underscores the elegant mechanism by which ATP synthase converts electrochemical potential energy into chemical energy, sustaining vital cellular functions.
| Characteristics | Values |
|---|---|
| Energy Source | Proton gradient (H⁺) across the inner mitochondrial membrane (in eukaryotes) or plasma membrane (in prokaryotes) |
| Process | Chemiosmosis |
| Mechanism | Proton flow through the ATP synthase enzyme complex drives the rotation of its central stalk, which in turn induces conformational changes in the catalytic sites to phosphorylate ADP to ATP |
| Location | Inner mitochondrial membrane (eukaryotes), thylakoid membrane (chloroplasts), plasma membrane (prokaryotes) |
| Substrates | ADP (adenosine diphosphate) and inorganic phosphate (Pi) |
| Product | ATP (adenosine triphosphate) |
| Driving Force | Electrochemical potential gradient of H⁺ ions |
| Origin of Proton Gradient | Oxidative phosphorylation (electron transport chain in mitochondria) or photophosphorylation (light-driven electron transport in chloroplasts) |
| Enzyme Structure | F₀ (membrane-embedded, proton channel) and F₁ (catalytic headpiece) subunits |
| Efficiency | Nearly 100% under optimal conditions |
| Regulation | Inhibited by ATP or high proton concentrations; activated by ADP and low proton concentrations |
| Biological Role | Primary mechanism for ATP production in cellular respiration and photosynthesis |
Explore related products






What You'll Learn
- Proton Motive Force: Proton gradient across membrane drives ATP synthesis in oxidative phosphorylation
- Electron Transport Chain: Transfers electrons, creating energy for proton pumping and ATP production
- Rotary Catalysis Mechanism: ATP synthase rotates, converting mechanical energy into chemical energy for ATP
- F1FO Complex Structure: Consists of F1 head and FO stalk, facilitating proton flow and synthesis
- Chemiosmosis Theory: Coupling of proton gradient to ATP synthesis via transmembrane potential
![]()
Proton Motive Force: Proton gradient across membrane drives ATP synthesis in oxidative phosphorylation
The proton motive force (PMF) is the driving power behind ATP synthesis in oxidative phosphorylation, a process fundamental to cellular energy production. This force arises from the electrochemical gradient of protons (H⁺ ions) across the inner mitochondrial membrane in eukaryotes or the plasma membrane in prokaryotes. As electrons move through the electron transport chain (ETC), they are transferred from high-energy donors like NADH and FADH₂ to lower-energy acceptors, ultimately reducing molecular oxygen to water. This electron flow pumps protons from the mitochondrial matrix (or cytoplasm in bacteria) into the intermembrane space, creating a concentration gradient and an electric potential difference. The PMF is the sum of these two components: the chemical potential (proton concentration difference) and the electrical potential (charge separation).
To visualize this process, imagine a dam holding back a reservoir of water. The protons, like water molecules, accumulate on one side of the membrane, creating a potential energy store. ATP synthase acts as a molecular turbine, embedded in the membrane and allowing protons to flow back into the matrix (or cytoplasm) through a channel. This flow drives the rotation of a central stalk within the enzyme, which in turn catalyzes the phosphorylation of ADP to ATP. Each proton translocated through ATP synthase supports the synthesis of approximately one ATP molecule, though the exact ratio can vary depending on factors like species and metabolic state. For instance, in mammalian mitochondria, the PMF typically drives a 2.5–3.0 H⁺/ATP ratio, reflecting the efficiency of the system.
One practical example of PMF’s role is in the uncoupling of oxidative phosphorylation by molecules like 2,4-dinitrophenol (DNP). DNP dissipates the proton gradient by allowing protons to re-enter the matrix without passing through ATP synthase, effectively "short-circuiting" the system. This uncoupling increases heat production at the expense of ATP synthesis, highlighting the critical dependence of ATP synthase on the PMF. Conversely, inhibitors like oligomycin block ATP synthase directly, preventing proton flow and halting ATP production, further underscoring the enzyme’s reliance on the gradient.
From a comparative perspective, the PMF is not exclusive to mitochondria; it is also central to energy transduction in chloroplasts during photosynthesis. Here, light-driven electron transport generates a proton gradient across the thylakoid membrane, which fuels ATP synthesis via CF₁FO ATP synthase. While the mechanisms differ—light energy drives proton pumping in photosynthesis, whereas redox reactions drive it in oxidative phosphorylation—the principle remains the same: a proton gradient powers ATP production. This universality highlights the PMF as a conserved and efficient mechanism for energy conversion across biological systems.
In practical terms, understanding the PMF has implications for metabolic disorders and therapeutic interventions. For example, mitochondrial diseases often involve defects in the ETC or ATP synthase, leading to reduced PMF and ATP depletion. Strategies to enhance PMF, such as calorie restriction or certain pharmacological agents, are being explored to mitigate these conditions. Additionally, in biotechnology, engineered systems like proton-pumping enzymes are used to generate PMF in synthetic cells, enabling ATP production in non-native environments. By harnessing the principles of the PMF, researchers aim to develop sustainable energy solutions and improve metabolic health, demonstrating its relevance beyond basic biology.
What Temperature Does Fuel Freeze? Understanding Diesel and Gasoline Freezing Points
You may want to see also
Explore related products



$80



![]()
Electron Transport Chain: Transfers electrons, creating energy for proton pumping and ATP production
The electron transport chain (ETC) is the powerhouse of cellular respiration, a complex series of protein complexes embedded in the inner mitochondrial membrane. Its primary function is to transfer electrons from high-energy molecules like NADH and FADH2, generated during earlier stages of respiration, to molecular oxygen (O2). This transfer is not a simple handoff; it’s a carefully orchestrated, stepwise process that harnesses the energy released at each step to pump protons (H⁺) across the membrane, creating an electrochemical gradient. This gradient, often referred to as the proton motive force, is the fuel that drives ATP synthase, the enzyme responsible for synthesizing ATP, the cell’s energy currency.
Consider the ETC as a molecular assembly line, where each station (Complex I to IV) performs a specific task. At Complex I, electrons from NADH are accepted and passed to ubiquinone (CoQ), a mobile electron carrier. This step is coupled with the pumping of four protons from the mitochondrial matrix to the intermembrane space. Similarly, Complex II receives electrons from FADH2, but pumps fewer protons. The electrons then move through the ubiquinone pool to Complex III, where they are transferred to cytochrome *c*, another mobile carrier, alongside the pumping of another four protons. Finally, at Complex IV, electrons reduce molecular oxygen to water, completing the chain. Each step is fine-tuned to maximize energy extraction, ensuring that the proton gradient is robust enough to power ATP synthase.
The efficiency of the ETC is remarkable, but it’s not without risks. Reactive oxygen species (ROS), such as superoxide and hydrogen peroxide, are inevitable byproducts of electron transport, particularly at Complex I and III. While low levels of ROS serve as signaling molecules, excessive production can damage cellular components, including DNA and lipids. Cells mitigate this through antioxidants like glutathione and enzymes like superoxide dismutase. Practical tips to support ETC function and minimize oxidative stress include consuming a diet rich in antioxidants (e.g., berries, nuts, and leafy greens) and maintaining a balanced intake of macronutrients to ensure a steady supply of NADH and FADH2 precursors.
Comparing the ETC to a hydroelectric dam highlights its elegance. Just as water flowing through turbines generates electricity, the flow of protons back into the matrix through ATP synthase drives the phosphorylation of ADP to ATP. The proton motive force, akin to the water pressure, determines the rate of ATP production. In humans, this process is so efficient that a single glucose molecule can yield up to 36 ATP molecules, though the actual number is often lower due to factors like substrate availability and metabolic state. For instance, during intense exercise, muscles rely heavily on the ETC to meet energy demands, emphasizing its critical role in sustaining life.
In conclusion, the electron transport chain is not merely a pathway for electron transfer but a sophisticated energy conversion system. By coupling electron flow with proton pumping, it creates the conditions necessary for ATP synthase to operate. Understanding its mechanics and vulnerabilities offers insights into optimizing cellular energy production and mitigating oxidative damage. Whether through dietary choices or lifestyle modifications, supporting the ETC’s function is key to maintaining overall health and vitality.
Ukraine Fuel Depot Attack: Unraveling the Truth Behind the Incident
You may want to see also
Explore related products






![]()
Rotary Catalysis Mechanism: ATP synthase rotates, converting mechanical energy into chemical energy for ATP
The rotary catalysis mechanism of ATP synthase is a marvel of molecular engineering, showcasing how nature harnesses mechanical energy to fuel life’s essential processes. At the heart of this mechanism is a rotating subunit, often likened to a molecular turbine, which spins at astonishing speeds—up to 150 revolutions per second in some bacteria. This rotation is driven by the proton gradient across the mitochondrial or bacterial membrane, created by the electron transport chain during oxidative phosphorylation. As protons flow through the enzyme’s rotor, they impart kinetic energy, transforming it into the chemical energy stored in ATP. This process is not just efficient; it’s nearly 100% effective in energy conversion, outperforming any human-made engine.
To visualize this mechanism, imagine a waterwheel powered by a flowing river. The river’s current (proton gradient) turns the wheel (rotor), which in turn drives a millstone (catalytic sites) to grind grain (synthesize ATP). In ATP synthase, the rotor’s movement triggers conformational changes in the catalytic sites, forcing ADP and inorganic phosphate to combine into ATP. Each revolution of the rotor produces 3 ATP molecules, a fixed ratio that underscores the precision of this molecular machine. This mechanism is so finely tuned that even slight disruptions, such as mutations in the enzyme’s structure, can halt ATP production, highlighting its critical role in cellular metabolism.
From a practical standpoint, understanding the rotary catalysis mechanism has profound implications for medicine and biotechnology. For instance, drugs targeting ATP synthase’s rotor could selectively inhibit energy production in pathogens like *Mycobacterium tuberculosis*, which rely heavily on this enzyme. Conversely, enhancing ATP synthase efficiency in human cells could mitigate energy deficits in aging or diseased tissues. Researchers are also exploring bioinspired designs, such as artificial molecular turbines, to replicate this mechanism for energy harvesting in nanodevices. By studying ATP synthase, scientists gain insights into optimizing energy conversion systems, both biological and synthetic.
Comparing ATP synthase to other energy-transducing enzymes reveals its uniqueness. Unlike static enzymes that rely on substrate binding for catalysis, ATP synthase couples mechanical motion to chemical synthesis, a strategy akin to a factory assembly line. This dynamic approach not only accelerates ATP production but also ensures spatial and temporal coordination with cellular energy demands. For example, in muscle cells under exertion, ATP synthase ramps up its rotation to meet the sudden surge in energy requirements. This adaptability underscores the elegance of the rotary catalysis mechanism, making it a cornerstone of bioenergetics.
In conclusion, the rotary catalysis mechanism of ATP synthase exemplifies nature’s ingenuity in converting energy forms. By rotating to synthesize ATP, this enzyme bridges the gap between physics and biochemistry, offering a blueprint for efficient energy conversion. Whether in drug development, bioengineering, or fundamental research, unraveling this mechanism opens doors to innovations that could revolutionize how we approach energy challenges in biology and beyond. Its study is not just an academic pursuit but a practical guide to harnessing the power of molecular machines.
Best Fuel Options for Hurricane Lamps: A Comprehensive Guide
You may want to see also
Explore related products






![]()
F1FO Complex Structure: Consists of F1 head and FO stalk, facilitating proton flow and synthesis
The F1FO ATP synthase complex is a molecular marvel, a rotary engine embedded in the membranes of mitochondria and bacteria, driving the synthesis of ATP, the energy currency of life. At its core lies a structural duality: the F1 head, a catalytic powerhouse, and the FO stalk, a proton-driven rotor. This elegant design harnesses the electrochemical gradient across the membrane, converting the flow of protons into mechanical energy, which in turn fuels ATP production.
Consider the F1 head as the factory floor where ATP is assembled. Composed of α and β subunits, it forms a hexameric structure with three catalytic sites. Each site undergoes a conformational change—open, loose, and tight—as protons flow through the FO stalk, inducing rotation. This rotation forces the β subunits to bind ADP and inorganic phosphate, catalyzing their union into ATP. The process is cyclical, with each 120-degree rotation synthesizing three ATP molecules, a testament to nature’s efficiency.
The FO stalk, by contrast, is the engine’s drive shaft. Embedded in the membrane, it consists of multiple subunits (e.g., *a*, *b*, and *c* in bacteria) that form a proton channel. Protons, driven by the membrane potential, pass through this channel, causing the *c*-ring to rotate. This rotation is transmitted to the central stalk, which connects the FO stalk to the F1 head, initiating the catalytic cycle. The number of *c* subunits determines the efficiency of this process; for instance, in *E. coli*, 10 *c* subunits require 3–4 protons per rotation, while in mitochondria, 8–15 *c* subunits optimize proton-to-ATP coupling.
To visualize this, imagine a waterwheel powered by a river’s flow. The FO stalk is the wheel, turning as protons pass through, while the F1 head is the mill, grinding out ATP with each rotation. This analogy underscores the complex’s reliance on proton flow, a process fueled by the electron transport chain in mitochondria or photosynthesis in chloroplasts. Without this gradient, the wheel stalls, and ATP synthesis halts.
Practical insights into this mechanism have led to targeted interventions in medicine and biotechnology. For example, inhibitors like oligomycin block the FO stalk, halting ATP production in pathogens while sparing host cells. Conversely, understanding the F1FO complex aids in designing proton-gradient-driven nanomachines for energy harvesting. Whether in a lab or a living cell, the F1FO complex remains a blueprint for converting energy gradients into usable work, a principle as applicable to synthetic biology as it is to cellular respiration.
Best Fuel Options for Your Honda Generator: A Comprehensive Guide
You may want to see also
Explore related products






![]()
Chemiosmosis Theory: Coupling of proton gradient to ATP synthesis via transmembrane potential
The chemiosmosis theory hinges on a simple yet profound concept: energy stored in the form of a proton gradient across a membrane drives ATP synthesis. This gradient, established by the active transport of protons (H⁺) out of the mitochondrial matrix or thylakoid lumen, creates an electrochemical potential difference. Think of it as a dam holding back water – the potential energy stored in the elevated water level is analogous to the energy stored in the proton gradient. When protons flow back across the membrane through ATP synthase, this energy is harnessed to phosphorylate ADP to ATP, much like the controlled release of water through a turbine generates electricity.
Mechanistically, ATP synthase acts as a molecular turbine, embedded in the inner mitochondrial membrane or thylakoid membrane. Its rotor subunit, F₀, is driven by the flow of protons down their concentration gradient. This rotational motion is coupled to the catalytic subunit, F₁, which binds and phosphorylates ADP. Each complete rotation of the F₀ subunit typically synthesizes 3 ATP molecules, showcasing the remarkable efficiency of this enzyme.
While the core principle of chemiosmosis is universal, its implementation differs between mitochondria and chloroplasts. In mitochondria, the proton gradient is generated by the electron transport chain (ETC) during oxidative phosphorylation. NADH and FADH₂, derived from the breakdown of glucose, donate electrons to the ETC, which pumps protons from the matrix to the intermembrane space. This gradient then fuels ATP synthase. In chloroplasts, light energy drives the formation of the proton gradient. Photosystem II splits water molecules, releasing electrons that ultimately reduce NADP⁺ to NADPH. Simultaneously, protons are pumped into the thylakoid lumen, creating a gradient that powers ATP synthase during photophosphorylation.
Practical implications of chemiosmosis extend to understanding metabolic disorders and developing therapeutic strategies. For instance, defects in the ETC complexes or ATP synthase itself can lead to mitochondrial diseases characterized by ATP depletion. Conversely, uncouplers like 2,4-dinitrophenol dissipate the proton gradient, preventing ATP synthesis and leading to increased metabolic rate – a principle exploited in some weight-loss supplements, albeit with significant risks.
In essence, the chemiosmosis theory reveals the elegant coupling of physical and chemical processes in energy transduction. By harnessing the power of proton gradients, cells efficiently convert energy from diverse sources – be it nutrients or sunlight – into the universal currency of ATP. This mechanism underscores the fundamental unity of bioenergetics across diverse organisms, from bacteria to humans.
Understanding Fuel Card Taxation: A Comprehensive Guide for Businesses
You may want to see also
Frequently asked questions
The primary source of energy for ATP synthase is the proton gradient (H⁺ gradient) established across the inner mitochondrial membrane or thylakoid membrane in chloroplasts. This gradient is created by the electron transport chain during oxidative phosphorylation or photosynthesis.
The proton gradient drives ATP synthase by allowing protons (H⁺ ions) to flow back into the mitochondrial matrix or stroma through the enzyme's rotor subunit. This flow of protons provides the mechanical energy needed to rotate the enzyme's central stalk, which in turn catalyzes the phosphorylation of ADP to ATP.
No, ATP synthase requires a proton gradient to function. Without the proton gradient, there is no driving force for the protons to flow through the enzyme, and thus, the mechanical energy needed to synthesize ATP is not generated. In some cases, ATP synthase can operate in reverse, hydrolyzing ATP to pump protons and maintain the gradient, but it cannot produce ATP without the gradient.































