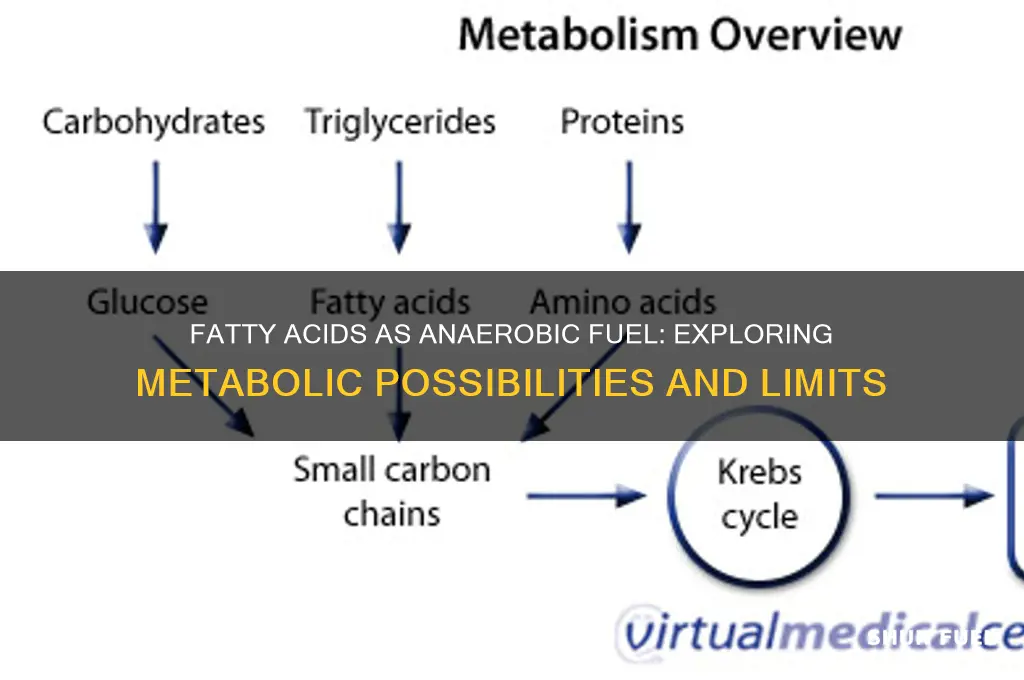
The utilization of fatty acids as a fuel source in metabolic processes is a well-established concept, primarily associated with aerobic respiration, where oxygen is required to break down fats efficiently. However, an intriguing question arises: can fatty acids be metabolized anaerobically, without the presence of oxygen? This topic delves into the potential of fatty acid oxidation in oxygen-depleted environments, exploring whether cells can harness energy from fats through alternative pathways. Understanding this process could have significant implications for various fields, including exercise physiology, where muscle cells might rely on fatty acid metabolism during intense, oxygen-limited activities, and in certain pathological conditions where oxygen availability is compromised. Investigating the anaerobic use of fatty acids may reveal novel insights into cellular energy production and its adaptability under different physiological and environmental conditions.
| Characteristics | Values |
|---|---|
| Can fatty acids be used as fuel anaerobically? | Yes, but with limitations. |
| Primary Anaerobic Fatty Acid Metabolism | Beta-oxidation in the cytoplasm, producing acetyl-CoA. |
| End Product of Anaerobic Fatty Acid Breakdown | Acetyl-CoA, which enters the ketogenesis pathway to produce ketone bodies (acetoacetate, beta-hydroxybutyrate, acetone). |
| Energy Yield Compared to Glucose | Lower ATP production per molecule compared to aerobic metabolism of fatty acids or glucose. |
| Tissues Capable of Anaerobic Fatty Acid Utilization | Primarily liver, as it's the main site of ketogenesis. Other tissues can utilize ketone bodies produced by the liver. |
| Physiological Conditions Favoring Anaerobic Fatty Acid Use | Prolonged fasting, starvation, intense exercise (when glycogen stores are depleted), and certain metabolic disorders. |
| Role of Ketone Bodies | Serve as alternative fuel source for brain, heart, and skeletal muscle during periods of low glucose availability. |
| Limitations | Ketogenesis is not a complete breakdown of fatty acids, and some carbon atoms are lost as CO2. Ketone bodies are less efficient energy sources than glucose. |
Explore related products






What You'll Learn
![]()
Fatty acid breakdown without oxygen
One key pathway for anaerobic fatty acid breakdown is anaerobic β-oxidation, which is distinct from the aerobic process. In this pathway, fatty acyl-CoA molecules are still broken down into acetyl-CoA units, but the electron transport chain, which typically relies on oxygen as the final electron acceptor, is bypassed. Instead, alternative electron acceptors such as nitrate, sulfate, or fumarate are used in microorganisms. For example, in some bacteria, the electrons derived from fatty acid oxidation are transferred to these acceptors through specialized reductases, allowing the process to continue without oxygen. This mechanism ensures that energy production from fatty acids can still occur in oxygen-limited environments.
In eukaryotic cells, anaerobic fatty acid breakdown is less common but can occur under specific conditions. For instance, during prolonged intense exercise, muscle cells may rely on anaerobic metabolism when oxygen delivery cannot meet the energy demand. However, fatty acids are not the preferred substrate for anaerobic energy production in these cells due to their slower oxidation rate compared to carbohydrates. Instead, glucose is primarily metabolized through glycolysis to produce ATP. Despite this, recent studies suggest that certain enzymes involved in fatty acid metabolism, such as carnitine palmitoyltransferase (CPT), may still play a role in facilitating limited fatty acid utilization even under anaerobic conditions.
Another aspect of anaerobic fatty acid breakdown involves the production of byproducts that differ from aerobic metabolism. In aerobic β-oxidation, the end products are CO₂ and water, but anaerobically, the process generates reduced molecules like succinate, acetate, or propionate, depending on the organism and the electron acceptor used. These byproducts can then enter other metabolic pathways, such as the citric acid cycle or fermentation processes, to generate additional ATP or serve as precursors for biosynthesis. This flexibility highlights the adaptability of fatty acid metabolism across diverse environmental conditions.
In summary, while aerobic β-oxidation is the primary pathway for fatty acid breakdown, anaerobic mechanisms exist to enable their utilization in oxygen-limited settings. These processes are particularly prominent in microorganisms but can also occur in eukaryotic cells under specific conditions. Understanding anaerobic fatty acid breakdown is crucial for fields such as microbiology, biochemistry, and physiology, as it sheds light on how organisms and cells adapt to survive and thrive in environments where oxygen is scarce.
Fuel Leaks and NC State Inspections: Will Your Vehicle Pass?
You may want to see also
Explore related products





![]()
Anaerobic metabolism pathways
Anaerobic metabolism refers to the process by which cells generate energy in the absence of oxygen. While fatty acids are primarily metabolized through aerobic pathways, such as beta-oxidation in the mitochondria, their utilization in anaerobic conditions is limited but not entirely impossible. Under anaerobic conditions, cells primarily rely on glycolysis and fermentation pathways to produce ATP. However, fatty acids cannot be directly used as a fuel source in these pathways due to their complex structure and the oxygen requirement for their breakdown. Beta-oxidation, the primary process for fatty acid breakdown, is strictly aerobic and occurs in the mitochondrial matrix, where molecular oxygen is essential for the final steps of the electron transport chain.
In anaerobic metabolism, glycolysis is the predominant pathway, where glucose is broken down into pyruvate, producing a small amount of ATP and NADH. Pyruvate is then converted into lactate in animals or ethanol in yeast through fermentation, allowing for the regeneration of NAD⁺, which is necessary for glycolysis to continue. Fatty acids do not enter this pathway because they cannot be converted into pyruvate or directly contribute to the glycolytic process. Instead, cells under anaerobic conditions prioritize carbohydrates as their primary fuel source due to the simplicity and efficiency of glycolysis in the absence of oxygen.
Despite the limitations, there is emerging evidence that certain anaerobic microorganisms can utilize fatty acids as a fuel source through modified metabolic pathways. For example, some bacteria and archaea employ anaerobic beta-oxidation or fatty acid activation mechanisms that do not require oxygen. These organisms often use alternative electron acceptors, such as sulfate or nitrate, to drive the oxidation of fatty acids. However, these processes are not considered true anaerobic metabolism in the context of higher eukaryotes, as they rely on specific microbial adaptations not present in animals or humans.
In higher organisms, the use of fatty acids as an anaerobic fuel is further constrained by the compartmentalization of metabolic pathways. Fatty acid oxidation occurs exclusively in the mitochondria, which is an aerobic environment. In contrast, anaerobic metabolism occurs in the cytoplasm, where glycolysis and fermentation take place. The physical separation of these compartments prevents fatty acids from being directly utilized in anaerobic conditions. Additionally, the transport of fatty acids into the mitochondria requires the carnitine shuttle system, which is energy-dependent and not functional under anaerobic conditions.
In summary, while fatty acids are a critical energy source in aerobic metabolism, their use in anaerobic conditions is highly restricted. Anaerobic metabolism pathways, such as glycolysis and fermentation, are designed to process carbohydrates, not fatty acids. Although certain microorganisms have evolved mechanisms to anaerobically metabolize fatty acids, these adaptations are not present in higher organisms. Thus, the use of fatty acids as fuel in anaerobic conditions remains largely infeasible in animals and humans, emphasizing the reliance on carbohydrates for energy production when oxygen is unavailable.
Can Fuel Tanks Be Repaired? Expert Insights and Repair Options
You may want to see also
Explore related products






![]()
Role of beta-oxidation in hypoxia
Beta-oxidation, the primary pathway for fatty acid metabolism, plays a critical role in energy production, particularly under conditions of hypoxia (low oxygen availability). While aerobic metabolism of fatty acids is well-understood, their utilization in anaerobic or hypoxic environments is less straightforward but still significant. Under normoxic conditions, fatty acids are broken down in the mitochondria through beta-oxidation, generating acetyl-CoA, which enters the citric acid cycle (TCA cycle) and ultimately produces ATP via oxidative phosphorylation. However, in hypoxia, oxidative phosphorylation is impaired due to limited oxygen, the terminal electron acceptor in the electron transport chain (ETC). Despite this, beta-oxidation can still occur, albeit with adaptations that allow cells to derive energy from fatty acids in the absence of sufficient oxygen.
In hypoxic conditions, such as those experienced by cells in ischemic tissues or tumors, beta-oxidation remains active but is uncoupled from the ETC. The acetyl-CoA produced from beta-oxidation can be redirected toward alternative metabolic pathways to generate ATP. One such pathway is anaerobic glycolysis, where pyruvate derived from glucose is converted to lactate, producing a small amount of ATP. However, fatty acids can also contribute to this process by providing acetyl-CoA, which can be converted to ketone bodies (e.g., acetoacetate and beta-hydroxybutyrate) in the liver or used directly in the TCA cycle in a truncated form. This allows cells to maintain energy production even when oxygen is scarce.
Another critical role of beta-oxidation in hypoxia is its ability to sustain membrane integrity and cellular function. Fatty acids are essential components of cell membranes, and their metabolism through beta-oxidation provides intermediates that can be used for lipid synthesis or repair. This is particularly important in hypoxic tissues, where cellular stress and damage are heightened. Additionally, beta-oxidation generates NADH and FADH2, which, although not fully utilized in the ETC under hypoxia, can still contribute to redox balance and support other metabolic processes, such as the synthesis of nucleotides and antioxidants.
The activation of beta-oxidation in hypoxia is regulated by key enzymes and transcription factors. For instance, peroxisome proliferator-activated receptors (PPARs), particularly PPAR-α, play a central role in upregulating genes involved in fatty acid oxidation. Under hypoxic conditions, the stabilization of hypoxia-inducible factor 1 (HIF-1) can also influence fatty acid metabolism by promoting the expression of enzymes that support beta-oxidation while inhibiting those that compete for acetyl-CoA in the TCA cycle. This regulatory interplay ensures that fatty acids are efficiently utilized as an energy source in oxygen-deprived environments.
In summary, beta-oxidation serves as a vital mechanism for fatty acid utilization in hypoxia, enabling cells to generate energy and maintain metabolic homeostasis despite limited oxygen availability. By redirecting acetyl-CoA toward alternative pathways and supporting membrane integrity, beta-oxidation ensures cellular survival in challenging conditions. Understanding its role in hypoxia not only sheds light on metabolic adaptations in diseases like cancer and ischemia but also highlights the versatility of fatty acid metabolism across diverse physiological states.
Can You Safely Split Fuel Nutrition Magnesium Tablets in Half?
You may want to see also
Explore related products






![]()
Fatty acids in anaerobic exercise
The utilization of fatty acids as a fuel source during exercise is a complex process that depends on the intensity and duration of the activity. While fatty acids are primarily associated with aerobic metabolism, their role in anaerobic exercise is a topic of interest. Anaerobic exercises, characterized by short bursts of high-intensity activity, typically rely on carbohydrates as the primary fuel source due to their rapid availability and efficiency in producing energy without oxygen. However, recent research suggests that fatty acids can still play a role in anaerobic conditions, albeit in a limited capacity.
During anaerobic exercise, the body’s energy demands exceed the oxygen supply, leading to the rapid breakdown of glycogen through glycolysis. This process produces ATP quickly but also results in the accumulation of lactate, which contributes to muscle fatigue. While fatty acid oxidation is generally an aerobic process requiring oxygen, emerging evidence indicates that fatty acids can contribute to energy production even in low-oxygen environments. This occurs through a process known as anaerobic β-oxidation, which is less efficient than aerobic oxidation but can still provide a small amount of ATP. Additionally, fatty acids can be used to regenerate NAD⁺, a coenzyme essential for glycolysis, thereby indirectly supporting anaerobic energy production.
The extent to which fatty acids are utilized during anaerobic exercise depends on several factors, including the individual’s fitness level, muscle fiber composition, and the availability of other fuel sources. For instance, individuals with higher mitochondrial density and greater capillary density in their muscles may have an enhanced capacity to oxidize fatty acids, even under anaerobic conditions. Furthermore, trained athletes often exhibit a greater reliance on fat oxidation during high-intensity exercise due to adaptations in muscle metabolism. However, it is important to note that fatty acids cannot fully replace carbohydrates as the primary fuel source during anaerobic activities.
Practical implications of fatty acid utilization in anaerobic exercise include dietary and training strategies to optimize performance. Consuming a diet rich in healthy fats can enhance the body’s ability to mobilize and oxidize fatty acids, potentially improving endurance during repeated anaerobic efforts. Additionally, incorporating high-intensity interval training (HIIT) can stimulate adaptations that increase fat oxidation capacity, even during short bursts of intense activity. While fatty acids may not be the dominant fuel source in anaerobic exercise, their contribution can help spare glycogen and delay fatigue, particularly in events requiring repeated sprints or power outputs.
In summary, while fatty acids are not the primary fuel source for anaerobic exercise, they can still contribute to energy production under these conditions. Through mechanisms such as anaerobic β-oxidation and NAD⁺ regeneration, fatty acids play a supportive role in sustaining high-intensity activity. Understanding this dynamic can inform training and nutritional strategies to enhance performance in anaerobic sports. Although carbohydrates remain essential for peak power output, optimizing fatty acid utilization can provide a valuable edge in endurance and recovery during repeated anaerobic efforts.
Leaky Truck Exhaust: A Hidden Cause of Fuel Starvation?
You may want to see also
Explore related products






![]()
Limitations of anaerobic fatty acid use
The utilization of fatty acids as a fuel source under anaerobic conditions presents several inherent limitations that constrain its efficiency and applicability. One primary limitation is the incomplete oxidation of fatty acids in the absence of oxygen. Unlike aerobic metabolism, where fatty acids are fully broken down into carbon dioxide and water via the citric acid cycle and oxidative phosphorylation, anaerobic conditions restrict the process to partial degradation. This typically results in the production of acetyl-CoA, which is further converted into molecules like acetoacetate, beta-hydroxybutyrate, and acetone—collectively known as ketone bodies. While ketone bodies can serve as alternative energy sources, their production is less energetically efficient compared to complete oxidation, yielding fewer ATP molecules per fatty acid molecule.
Another significant limitation is the accumulation of byproducts that can disrupt cellular homeostasis. Anaerobic fatty acid metabolism, particularly in prolonged or intense conditions, leads to the buildup of ketone bodies and other intermediates. High levels of ketones can cause metabolic acidosis, a condition where the blood pH drops due to excess acid, impairing cellular function and potentially leading to organ damage. Additionally, the liver, which is central to ketone body production, can become overwhelmed, further limiting the sustainability of this metabolic pathway.
The reliance on anaerobic fatty acid use is also constrained by the limited capacity of tissues to utilize ketone bodies as fuel. While certain tissues, such as the brain and skeletal muscle, can metabolize ketones, others, like the erythrocytes (red blood cells), lack the necessary enzymes to do so. This tissue-specific limitation reduces the overall effectiveness of fatty acids as a universal anaerobic fuel source. Furthermore, the transition to ketone body utilization requires adaptive changes in cellular metabolism, which may not occur rapidly enough to meet immediate energy demands during anaerobic conditions.
Enzymatic and transport limitations further hinder anaerobic fatty acid use. The transport of fatty acids into the mitochondria, where beta-oxidation occurs, relies on specific carnitine-dependent mechanisms. Under anaerobic conditions, the efficiency of these transport systems may be compromised, reducing the availability of fatty acids for metabolism. Additionally, the enzymes involved in beta-oxidation and ketogenesis may become rate-limiting, particularly in the absence of oxygen, which affects the electron transport chain and overall metabolic flux.
Lastly, the sustainability of anaerobic fatty acid use is limited by the body's finite fatty acid stores and the challenges of replenishing them under anaerobic conditions. Unlike glucose, which can be rapidly replenished through glycogenolysis or gluconeogenesis, fatty acids are primarily derived from adipose tissue and dietary intake. Prolonged reliance on fatty acids under anaerobic conditions can deplete these stores, leading to energy deficits and metabolic stress. This limitation is particularly relevant in scenarios such as intense exercise or hypoxic environments, where energy demands are high but oxygen availability is low.
In summary, while fatty acids can be utilized as a fuel source under anaerobic conditions, their use is constrained by incomplete oxidation, byproduct accumulation, tissue-specific limitations, enzymatic inefficiencies, and finite storage capacity. These limitations highlight the challenges of relying on fatty acids as a primary energy source in oxygen-deprived settings and underscore the importance of aerobic metabolism for efficient fatty acid utilization.
Can a Faulty Fuel Damper Reduce Engine Power?
You may want to see also
Frequently asked questions
Yes, fatty acids can be partially broken down through anaerobic processes like beta-oxidation, but complete oxidation requires oxygen.
During anaerobic metabolism, fatty acids undergo incomplete breakdown, producing acetyl-CoA, which can enter the citric acid cycle, but the process is limited due to the lack of oxygen.
No, carbohydrates (glycogen) are the primary fuel source for anaerobic exercise due to their faster breakdown and energy production without oxygen.
Fatty acids can contribute to ATP production anaerobically through limited beta-oxidation and ketogenesis, but the yield is significantly lower compared to aerobic conditions.
Fatty acid metabolism is less efficient anaerobically because the electron transport chain, which requires oxygen, is bypassed, limiting the amount of ATP that can be generated.































