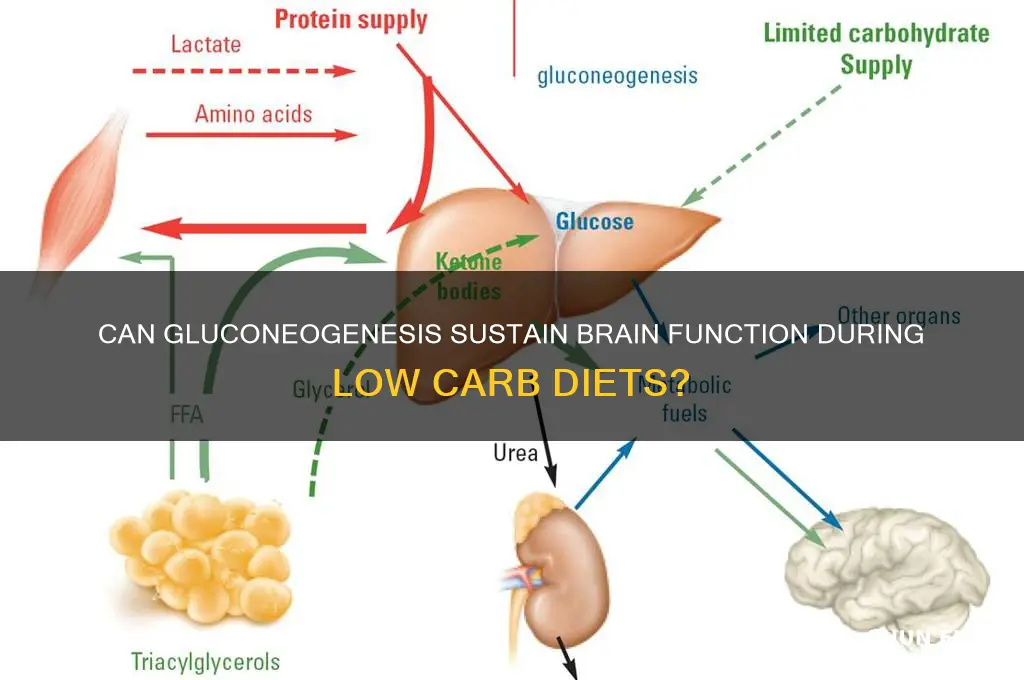
Gluconeogenesis, the process by which the body synthesizes glucose from non-carbohydrate precursors like amino acids and glycerol, plays a critical role in maintaining blood glucose levels during fasting or low-carbohydrate intake. While the brain is highly dependent on glucose as its primary energy source, it can only utilize glucose derived from the bloodstream, as it lacks the necessary enzymes for gluconeogenesis. This raises the question: can gluconeogenesis effectively fuel the brain? Although gluconeogenesis ensures a steady supply of glucose to the bloodstream, the brain’s reliance on this process is indirect, as it depends on the liver to produce and release glucose. During prolonged fasting or intense metabolic demands, gluconeogenesis becomes essential to prevent hypoglycemia and sustain brain function, highlighting its indirect yet vital role in supporting cerebral energy needs.
| Characteristics | Values |
|---|---|
| Primary Brain Fuel | Glucose is the primary and preferred energy source for the brain under normal conditions. |
| Gluconeogenesis Role | Gluconeogenesis (GNG) can produce glucose from non-carbohydrate precursors (e.g., lactate, glycerol, amino acids) when dietary carbohydrate intake is low or during fasting. |
| Brain's Glucose Dependency | The brain typically consumes ~120 g of glucose daily, accounting for ~20% of total body glucose utilization. |
| GNG Contribution to Brain Fuel | During prolonged fasting or low-carb diets, GNG can supply up to 50-75% of the brain's glucose needs, with the remainder coming from glycogenolysis or ketone bodies. |
| Ketone Bodies as Alternative Fuel | In states of prolonged fasting or ketosis, the brain can adapt to use ketone bodies (β-hydroxybutyrate and acetoacetate) for up to 70% of its energy needs, reducing reliance on GNG-derived glucose. |
| Amino Acid Utilization | GNG uses amino acids (e.g., alanine, glutamine) as substrates, which can limit protein availability for other bodily functions if overutilized. |
| Lactate as Substrate | Lactate from muscle glycogen breakdown can be a significant substrate for GNG, contributing to brain glucose supply during exercise or fasting. |
| Glycerol Contribution | Glycerol from triglyceride breakdown can also serve as a substrate for GNG, particularly during prolonged fasting. |
| Limitations of GNG | GNG has a limited capacity to fully replace dietary glucose intake for the brain, especially in the absence of ketone body utilization. |
| Adaptations in Fasting | Prolonged fasting enhances GNG efficiency and increases reliance on ketone bodies, allowing the brain to function with reduced glucose input. |
| Clinical Relevance | Disorders of GNG (e.g., glycogen storage diseases) can impair brain glucose supply, highlighting its importance in metabolic homeostasis. |
Explore related products






What You'll Learn
![]()
Glucose Demand in Brain Function
The brain is an exceptionally energy-demanding organ, accounting for approximately 20% of the body's total glucose utilization despite representing only about 2% of body weight. This high demand is primarily due to the brain's reliance on glucose as its primary fuel source under normal physiological conditions. Glucose is essential for maintaining neuronal function, synaptic transmission, and the integrity of the blood-brain barrier. Unlike other tissues, the brain has limited capacity to utilize alternative energy sources such as fatty acids, which cannot cross the blood-brain barrier effectively. This makes glucose supply critical for sustaining cognitive processes, consciousness, and overall brain health.
Under typical circumstances, the brain's glucose requirements are met through systemic blood glucose levels, which are tightly regulated by hormonal mechanisms involving insulin and glucagon. However, during periods of fasting, hypoglycemia, or intense mental activity, the brain's glucose demand must be met through alternative pathways. This raises the question of whether gluconeogenesis, the process by which glucose is synthesized from non-carbohydrate precursors such as amino acids and glycerol, can contribute to fueling the brain. While gluconeogenesis primarily occurs in the liver and, to a lesser extent, the kidneys, the glucose produced is released into the bloodstream and becomes available for brain uptake.
Gluconeogenesis plays a crucial role in maintaining blood glucose levels during prolonged fasting or low-carbohydrate diets, thereby indirectly supporting brain function. However, the extent to which gluconeogenesis directly fuels the brain is limited by the brain's high glucose consumption rate and the relatively slow pace of gluconeogenic glucose production. Studies suggest that gluconeogenesis can provide approximately 50-75% of the glucose needed during fasting states, with the remainder derived from glycogenolysis (breakdown of liver glycogen). Despite this, the brain's reliance on a continuous glucose supply means that gluconeogenesis alone cannot fully meet its energy demands, especially during periods of increased metabolic activity.
Another factor to consider is the brain's ability to adapt to reduced glucose availability through mechanisms such as increased ketone body utilization. During prolonged fasting or ketogenic states, the brain can derive up to 70% of its energy from ketones, reducing its dependence on glucose. However, this adaptation is not immediate and requires several days of carbohydrate restriction. In the interim, gluconeogenesis remains a vital process for preventing hypoglycemia and ensuring that the brain receives sufficient glucose to maintain function. Thus, while gluconeogenesis is not the primary fuel source for the brain, it serves as a critical supplementary mechanism to sustain glucose levels and support brain function in the absence of dietary carbohydrates.
In summary, the brain's glucose demand is both substantial and non-negotiable, making it heavily reliant on systemic glucose availability. While gluconeogenesis is essential for maintaining blood glucose levels during fasting or low-carbohydrate conditions, it cannot fully replace dietary glucose as the brain's primary fuel source. The brain's limited ability to utilize alternative fuels, combined with the relatively slow rate of gluconeogenic glucose production, underscores the importance of dietary carbohydrate intake or glycogen stores in meeting its energy needs. Understanding the interplay between gluconeogenesis, glucose supply, and brain function highlights the complexity of metabolic adaptations and the brain's unique metabolic requirements.
Can a Faulty Fuel Injector Trigger Your Check Engine Light?
You may want to see also
Explore related products






![]()
Alternative Energy Sources for Brain
The brain is a highly energy-demanding organ, typically relying on glucose as its primary fuel source. However, under certain conditions, such as prolonged fasting or low carbohydrate intake, the brain must adapt to alternative energy sources. Gluconeogenesis, the process of synthesizing glucose from non-carbohydrate precursors like amino acids and glycerol, plays a crucial role in maintaining blood glucose levels. While gluconeogenesis ensures a steady supply of glucose to the brain during fasting, it is not the only mechanism by which the brain can sustain its energy needs. Research indicates that the brain can utilize ketone bodies, derived from the breakdown of fats in the liver, as a significant alternative fuel source. During prolonged fasting or ketogenic diets, ketone bodies such as beta-hydroxybutyrate and acetoacetate can provide up to 70% of the brain's energy requirements, reducing its dependence on glucose.
Another alternative energy source for the brain is lactate, which is produced by muscles and other tissues during anaerobic metabolism. Under conditions of intense physical activity or hypoxia, lactate can be taken up by the brain and used as a fuel source. Studies suggest that lactate may account for a substantial portion of the brain's energy needs during exercise, highlighting its role as a flexible and readily available energy substrate. Additionally, lactate can enhance neuronal metabolism and support cognitive function, making it a valuable alternative to glucose.
Emerging research also points to the potential role of medium-chain triglycerides (MCTs) in fueling the brain. Unlike long-chain fatty acids, MCTs can cross the blood-brain barrier and be directly metabolized by brain cells. MCTs are rapidly converted into ketones, providing a quick and efficient energy source for the brain. This has led to their use in therapeutic applications, such as managing epilepsy or cognitive decline in conditions like Alzheimer's disease, where glucose metabolism is impaired.
Furthermore, amino acids can serve as an indirect energy source for the brain through gluconeogenesis, but they also play a role in neurotransmitter synthesis and neuronal function. While the brain cannot directly use amino acids for energy, their contribution to maintaining overall metabolic homeostasis is vital. For instance, branched-chain amino acids (BCAAs) can be oxidized in muscles, producing energy and freeing up glucose for the brain.
In summary, while gluconeogenesis is essential for maintaining glucose levels during fasting, the brain is remarkably adaptable and can utilize ketone bodies, lactate, MCTs, and indirectly, amino acids, as alternative energy sources. Understanding these pathways not only sheds light on the brain's metabolic flexibility but also opens avenues for therapeutic interventions in conditions where glucose metabolism is compromised. By leveraging these alternative fuels, it is possible to support brain function in diverse physiological and pathological states.
Boosting Engine Power: Can Fuel Pumps Enhance Performance?
You may want to see also
Explore related products






![]()
Role of Ketone Bodies
The brain is a highly energy-demanding organ, typically relying on glucose as its primary fuel source. However, during states of low glucose availability, such as prolonged fasting, starvation, or a low-carbohydrate ketogenic diet, the brain must adapt to alternative energy sources. While gluconeogenesis—the process of synthesizing glucose from non-carbohydrate precursors—can provide some glucose, it is insufficient to meet the brain's substantial energy demands. This is where ketone bodies play a crucial role. Ketone bodies, including acetoacetate, β-hydroxybutyrate, and acetone, are produced in the liver from fatty acids during prolonged fasting or carbohydrate restriction. They serve as an efficient alternative fuel for the brain, compensating for the reduced glucose supply.
Ketone bodies are particularly important because they can cross the blood-brain barrier and are readily taken up by brain cells. Unlike other fatty acids, which cannot directly fuel the brain, ketone bodies can enter the mitochondria and undergo oxidation to produce ATP. This process becomes essential when glucose levels are low, as the brain cannot store significant amounts of glucose. Studies have shown that during ketosis, ketone bodies can provide up to 70% of the brain's energy needs, significantly reducing its reliance on glucose. This metabolic flexibility ensures that the brain remains functional even in the absence of adequate carbohydrate intake.
The utilization of ketone bodies by the brain is facilitated by specific enzymes and transporters. Monocarboxylate transporters (MCTs), particularly MCT1, are responsible for transporting ketone bodies across the blood-brain barrier and into neurons. Once inside the cell, ketone bodies are converted into acetyl-CoA, which enters the citric acid cycle to generate ATP. This pathway is highly efficient and produces more ATP per molecule compared to glucose metabolism. Additionally, ketone metabolism generates fewer reactive oxygen species (ROS), potentially offering neuroprotective benefits by reducing oxidative stress.
Another critical aspect of ketone bodies is their role in maintaining cognitive function during glucose deprivation. Research has demonstrated that ketone bodies can preserve neuronal integrity and support synaptic function, even under conditions of metabolic stress. This is particularly relevant in pathological states such as epilepsy, Alzheimer's disease, and traumatic brain injury, where ketone bodies have been shown to improve cognitive outcomes. For example, the ketogenic diet, which elevates ketone body production, is a well-established treatment for drug-resistant epilepsy, highlighting the therapeutic potential of ketone bodies in neurological disorders.
In summary, while gluconeogenesis can contribute to glucose supply for the brain, it is ketone bodies that play the dominant role in fueling the brain during periods of carbohydrate restriction or fasting. Their ability to cross the blood-brain barrier, efficiently produce ATP, and provide neuroprotective effects makes them indispensable for brain function under metabolic stress. Understanding the role of ketone bodies not only sheds light on the brain's adaptability but also opens avenues for therapeutic interventions in various neurological conditions.
Can Fuel Injectors Be Repaired, Replaced, or Cleaned for Optimal Performance?
You may want to see also
Explore related products






![]()
Limitations of Gluconeogenesis
Gluconeogenesis, the process by which the body synthesizes glucose from non-carbohydrate precursors, is a vital metabolic pathway, particularly during periods of fasting or low carbohydrate intake. However, when considering whether gluconeogenesis can fully fuel the brain, several limitations become apparent. The brain is a highly glucose-dependent organ, typically consuming about 20% of the body’s glucose supply under normal conditions. While gluconeogenesis can produce glucose, its capacity is finite and influenced by substrate availability, primarily lactate, glycerol, and certain amino acids. During prolonged fasting, the depletion of these substrates limits the rate of gluconeogenesis, making it insufficient to meet the brain’s constant energy demands.
Another limitation lies in the brain’s adaptability to alternative fuel sources. Although the brain can utilize ketone bodies (derived from fatty acid metabolism) as an energy source during prolonged fasting or ketogenic states, this transition is not immediate. The brain still requires a baseline amount of glucose, which gluconeogenesis alone cannot always provide in sufficient quantities. Additionally, not all brain regions can efficiently use ketones, further emphasizing the brain’s reliance on glucose. This partial dependence on glucose highlights a critical limitation of gluconeogenesis in fully sustaining brain function.
The efficiency and energy cost of gluconeogenesis also pose significant constraints. Gluconeogenesis is an energy-intensive process, requiring six ATP molecules to produce one molecule of glucose from lactate or glycerol. This inefficiency becomes particularly problematic during states of energy deficit, such as starvation or intense physical activity, where ATP reserves are already compromised. The body prioritizes conserving energy, often reducing the rate of gluconeogenesis to preserve ATP for more critical functions, thereby limiting glucose availability for the brain.
Furthermore, the reliance of gluconeogenesis on amino acids, particularly those derived from muscle protein breakdown, introduces long-term limitations. Prolonged use of amino acids for gluconeogenesis can lead to muscle wasting and metabolic imbalances, as the body sacrifices lean tissue to maintain blood glucose levels. This is unsustainable and detrimental to overall health, underscoring the impracticality of gluconeogenesis as a long-term solution for brain fuel.
Lastly, individual variability in metabolic efficiency and health conditions can further restrict the effectiveness of gluconeogenesis. Factors such as insulin resistance, liver dysfunction, or genetic disorders can impair the gluconeogenic pathway, reducing its ability to produce adequate glucose for the brain. In such cases, even under optimal conditions, gluconeogenesis may fall short of meeting the brain’s glucose requirements, necessitating external glucose intake or medical intervention.
In summary, while gluconeogenesis plays a crucial role in maintaining blood glucose levels, its limitations in substrate availability, energy efficiency, long-term sustainability, and individual variability make it insufficient to fully fuel the brain, particularly under prolonged or stressful conditions. The brain’s unique metabolic demands necessitate a multifaceted approach to energy supply, combining gluconeogenesis with other pathways like ketogenesis and dietary glucose intake.
Rebuilding Fuel Injectors: Cost-Effective Solution or Waste of Time?
You may want to see also
Explore related products






![]()
Brain’s Metabolic Flexibility
The brain is an exceptionally energy-demanding organ, accounting for approximately 20% of the body’s total energy expenditure despite representing only 2% of body weight. To meet this high demand, the brain exhibits remarkable metabolic flexibility, the ability to adapt its fuel utilization based on availability. While glucose is traditionally considered the brain’s primary energy source, emerging research highlights its capacity to utilize alternative fuels, particularly under conditions of glucose scarcity. This adaptability is crucial for survival, as it ensures continuous energy supply to maintain cognitive function and neuronal integrity.
Under normal circumstances, the brain relies heavily on glucose, which is transported across the blood-brain barrier via glucose transporters (GLUT1). However, during states of fasting, prolonged exercise, or low carbohydrate intake, blood glucose levels decline, prompting the body to engage in gluconeogenesis—the synthesis of glucose from non-carbohydrate precursors such as lactate, glycerol, and amino acids. While gluconeogenesis primarily occurs in the liver and kidneys, the brain can indirectly benefit from this process. Lactate, produced by muscles during exercise or by astrocytes in the brain, can be taken up by neurons and converted back to pyruvate for oxidative metabolism, effectively bypassing the need for direct glucose supply.
Despite gluconeogenesis not directly fueling the brain, its role in maintaining systemic glucose levels is vital for brain function. During prolonged fasting, gluconeogenesis ensures a steady supply of glucose to the brain, preventing hypoglycemia and supporting neuronal activity. Additionally, ketone bodies, produced in the liver from fatty acids during prolonged fasting or ketogenic diets, emerge as a significant alternative fuel for the brain. Neurons can efficiently metabolize ketones, particularly acetoacetate and β-hydroxybutyrate, via the tricarboxylic acid (TCA) cycle, reducing reliance on glucose. This metabolic shift underscores the brain’s flexibility in utilizing available energy substrates.
The interplay between gluconeogenesis, ketogenesis, and brain metabolism highlights the brain’s ability to prioritize energy homeostasis. While gluconeogenesis does not directly fuel the brain, it sustains glucose levels, allowing the brain to function until alternative fuels like ketones become dominant. This adaptive mechanism is particularly critical in pathological conditions such as diabetes, where impaired glucose regulation necessitates reliance on other energy sources. Furthermore, astrocytes play a pivotal role in this metabolic flexibility by storing glycogen, which can be broken down to lactate and shuttled to neurons during energy crises.
In summary, the brain’s metabolic flexibility is a testament to its evolutionary design to ensure survival under varying nutritional states. While gluconeogenesis does not directly fuel the brain, it indirectly supports brain function by maintaining systemic glucose levels. The brain’s ability to switch between glucose, lactate, and ketones as energy sources demonstrates its resilience and adaptability. Understanding these mechanisms not only sheds light on normal brain physiology but also offers insights into therapeutic strategies for metabolic disorders and neurological conditions where energy metabolism is compromised.
Using Fuel Stabilizer in a 1966 Corvette: What You Need to Know
You may want to see also
Frequently asked questions
Yes, gluconeogenesis can supply glucose to the brain during fasting or low-carb diets. The brain relies on glucose as its primary fuel, and when dietary carbohydrates are insufficient, gluconeogenesis in the liver converts non-carbohydrate substrates like amino acids and glycerol into glucose to meet the brain's energy demands.
Gluconeogenesis can provide a significant portion of the brain's glucose requirements, but not all. Under normal conditions, the brain uses about 120 grams of glucose daily. During prolonged fasting, gluconeogenesis can produce around 80-100 grams of glucose per day, with the remaining energy needs being met by ketone bodies, which the brain can use as an alternative fuel source.
No, gluconeogenesis cannot fully replace dietary glucose for brain function, especially in the absence of ketone bodies. While it can produce glucose, the brain still requires a steady supply of glucose or ketones to function optimally. Prolonged reliance on gluconeogenesis alone, without adequate dietary intake or ketone production, can lead to energy deficits and impaired brain function.






























